WikiDer > Гнездо (структурный мотив белка)
В Гнездо это тип структурный мотив белка. Это небольшая повторяющаяся функция связывания анионов обоих белки и пептиды.[1][2][3][4][5][6][7][8][9] Каждый состоит из атомов основной цепи трех последовательных аминокислота остатки. Группы NH основной цепи связывают анионы, в то время как атомы боковой цепи часто не участвуют. Пролин в остатках отсутствуют группы NH, поэтому они редко встречаются в гнездах. В среднем около одного из 12 аминокислотных остатков в белках приходится на гнездо.
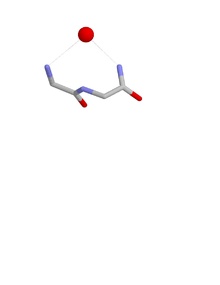
Конформации гнезда
Конформация гнезда такова, что группы NH первого и третьего аминокислотных остатков могут быть водородная связь к отрицательно заряженному или частично отрицательно заряженному атому, часто к атому кислорода. NH второго остатка также может быть связан водородной связью с тем же атомом, но обычно указывает несколько в сторону. Эти атомы главной цепи образуют углубление, называемое гнездом, в которое помещается анионный атом. Такие анионные атомы иногда называют яйцами, и более одного яйца может быть связано с гнездом. В оксианион отверстие кишечника сериновые протеазы это функциональный пример гнезда. Другой происходит на дне глубокой полости в антибиотик пептид ванкомицин который связывает ключ карбоксилат группа, используемая на заключительных этапах стенка бактериальной клетки синтез, тем самым предотвращая размножение бактериальных клеток.
Гнезда определяются конформацией атомов основной цепи, а именно phi, psi двугранные углы первых двух аминокислот в гнезде. Для типичного (RL) гнезда phiя= -90 °; psiя= 0 °; фия + 1= 80 °; psiя + 1=20°.
Гнезда различаются по степени вогнутости. У некоторых так мало, что теряется вогнутость; эти пептиды часто связывают катионы через свои CO-группы основной цепи, а не анионы через свои NH-группы. Фильтр специфичности калиевый канал[10] и водный канал аквапорин демонстрируют эту более линейную конформацию, в которой карбонильные группы используются белками для транспортировки молекул через мембраны. Эта почти линейная конформация также встречается в цепи альфа-лист[11][12][13]
Составные гнезда
Если два гнезда перекрываются так, что остаток i + 1 первого гнезда является остатком i второго гнезда, формируется составное гнездо. Здесь четыре группы NH вместо трех. Если три гнезда перекрываются, так что остатки i + 1 и i + 2 первого гнезда являются остатком i второго и третьего гнезда, формируется более широкое составное гнездо с пятью группами NH и так далее. Атомы основной цепи образуют часть неполного кольца, причем все группы NH ориентированы примерно к центру кольца. Поскольку их вогнутости часто шире, чем у простых гнезд, составные гнезда обычно используются белками для связывания многоатомных анионов, таких как фосфаты, как в P-петля или же Мотивы Walker, И в железо-серные кластеры. Синтезированный пептид Ser-Gly-Ala-Gly-Lys-Thr, сконструированный как минимальный пептид. P-петля, было показано, что он сильно связывает неорганический фосфат при нейтральном pH. [14].
Виды гнезд
Простые гнезда бывают двух видов, называемых RL и LR, в зависимости от знака углов фи первых двух остатков гнезда. Остатки R имеют отрицательные значения phi (как в правой альфа-спирали), а остатки L имеют положительные значения phi (как в левой альфа спираль). Восемьдесят процентов гнезд являются RL и 20% - LR. Когда два гнезда перекрываются, они могут быть RLR или LRL. Когда три гнезда перекрываются, они могут быть RLRL или LRLR и так далее.
Каждый Петля Шельмана включает гнездо RL в последних трех из шести остатков. Гнездо связывает карбонил атомы кислорода, предшествующие ему по порядку.
Номер антитело белки имеют гнезда RLR внутри шпилечных петель их CDR H-цепи (регионы, определяющие комплементарность), связанный с боковой цепью карбоксилата. Они были разработаны для получения моноклональных антител, содержащих гнезда, специфичных для белков с фосфорилированными серины и треонины.[15]
Наиболее PDZ домены имеют гнездо RL в начале первой бета-цепи с функцией распознавания карбоксилатной группы на С-конце пептида или белкового лиганда домена.[16]
Рекомендации
- ^ Watson, JD; Милнер-Уайт (2002). «Новый сайт связывания анионов основной цепи в белках: гнездо. Конкретная комбинация значений phi, psi в последовательных остатках дает начало сайтам связывания анионов, которые обычно встречаются и часто находятся в функционально важных областях». Журнал молекулярной биологии. 315 (2): 171–182. Дои:10.1006 / jmbi.2001.5227. PMID 11779237.
- ^ Pal, D; Зунель (2002). «Новые принципы построения белка: гнезда, яйца и что дальше?». Angew Chem Int Ed. 41 (24): 4663–4665. Дои:10.1002 / anie.200290009. PMID 12481319.
- ^ Милнер-Уайт, EJ; Ниссинк (2004). «Повторяющиеся анион-связывающие мотивы основной цепи в коротких полипептидах: гнезда». Acta Crystallographica Раздел D. D60 (11): 1935–1942. Дои:10.1107 / s0907444904021390. PMID 15502299.
- ^ Паевски, Р; Фердани (2005). «Катионная зависимость комплексообразования хлорид-иона с помощью разомкнутых рецепторных молекул в растворе хлороформа». Журнал Американского химического общества. 127 (51): 18281–18295. Дои:10.1021 / ja0558894. PMID 16366583.
- ^ Беркессель, А; Кох (2006). «Асимметричное эпоксидирование енона твердофазными связанными пептидами: дополнительные доказательства спиральности катализатора и каталитической активности отдельных цепей». Биополимеры. 84 (1): 90–96. Дои:10.1002 / bip.20413. PMID 16283656.
- ^ Милнер-Уайт, EJ; Рассел (2006). «Предсказание конформации белков и пептидов на ранней стадии эволюции». Биология Директ. 3: 3. Дои:10.1186/1745-6150-3-3. ЧВК 2241844. PMID 18226248.
- ^ Watson, JD; Ласковский (2005). «ProFunc: сервер для прогнозирования функции белка по трехмерной структуре». Исследования нуклеиновых кислот. 33 (Веб-сервер): W89 – W93. Дои:10.1093 / нар / gki414. ЧВК 1160175. PMID 15980588.
- ^ Лэнгтон, MJ; Serpell CJ; Пивной ПД (2016). «Распознавание анионов в воде: последние достижения с надмолекулярной и макромолекулярной точки зрения». Angewandte Chemie International Edition. 55 (6): 1974–1987. Дои:10.1002 / anie.201506589. ЧВК 4755225. PMID 26612067.
- ^ Cremer, P; Flood AS; Гибб BC; Мобли Д.Л. (2018). «Совместные пути к прояснению мутных вод водной надмолекулярной химии». Химия природы. 10 (1): 8–16. Дои:10.1038 / nchem.2894. PMID 29256514.
- ^ Watson, JD; Милнер-Уайт (2002). «Конформации полипептидных цепей, в которых части основной цепи последовательных остатков являются энантиомерами. Их наличие в катионных и анион-связывающих областях белков». Журнал молекулярной биологии. 315 (15): 183–191. Дои:10.1006 / jmbi.2001.5228. PMID 11779238.
- ^ Милнер-Уайт, EJ; Уотсон (2006). «Образование амилоида может включать взаимное преобразование альфа- в бета-листы посредством переворачивания пептидной плоскости». Структура. 14 (9): 1369–1376. Дои:10.1016 / j.str.2006.06.016. PMID 16962968.
- ^ Hayward, S; Милнер-Уайт (2008). «Геометрия α-листа: последствия для его возможной функции в качестве предшественника амилоида в белках». Белки. 71 (1): 415–425. Дои:10.1002 / prot.21717. PMID 17957773.
- ^ Hayward, S; Милнер-Уайт (2011). «Моделирование перехода β- в α-лист приводит к скрученному листу для антипараллельных и α-нанотрубке для параллельных нитей: последствия для образования амилоида». Белки. 79 (11): 3193–3207. Дои:10.1002 / prot.23154. PMID 21989939.
- ^ Бьянки, А; Георгий А; Ruzza P; Тониоло С. (2013). «Показано, что синтетический гексапептид, напоминающий белковое гнездо P-петли, связывает неорганический фосфат». Белки. 80 (5): 1418–1424. Дои:10.1002 / prot.24038. PMID 22275093.
- ^ Koerber, JT; Томсен Н.Д .; Ханниган БТ; DeGrado WF; Уэллс Дж. А. (2013). «Вдохновленный природой дизайн каркасов специфических антител». Природа Биотехнологии. 31 (10): 916–921. Дои:10.1038 / nbt.2672. ЧВК 3795957. PMID 23955275.
- ^ Ли, HJ; Чжэн Дж.Дж. (2010). «Домены PDZ и их партнеры по связыванию: специфика структуры и модификация». Сотовая связь и сигнализация. 8: 8. Дои:10.1186 / 1478-811x-8-8. ЧВК 2891790. PMID 20509869.
внешняя ссылка
- ^ Лидер, ДП; Милнер-Уайт (2009). «Мотивированные белки: веб-приложение для изучения небольших трехмерных белковых мотивов». BMC Биоинформатика. 10 (1): 60. Дои:10.1186/1471-2105-10-60. ЧВК 2651126. PMID 19210785.
- ^ Головин А; Хенрик (2008). «MSDmotif: изучение сайтов и мотивов белков». BMC Биоинформатика. 9 (1): 312. Дои:10.1186/1471-2105-9-312. ЧВК 2491636. PMID 18637174.