WikiDer > Cosesaurus
| Cosesaurus | |
|---|---|
 | |
| Реконструкция жизни Cosesaurus aviceps | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Учебный класс: | Рептилии |
| Семья: | †Tanystropheidae |
| Род: | †Cosesaurus Элленбергер и де Вильяльта 1974 |
| Разновидность: | †С. aviceps |
| Биномиальное имя | |
| †Cosesaurus aviceps Элленбергер и де Вильяльта 1974 | |
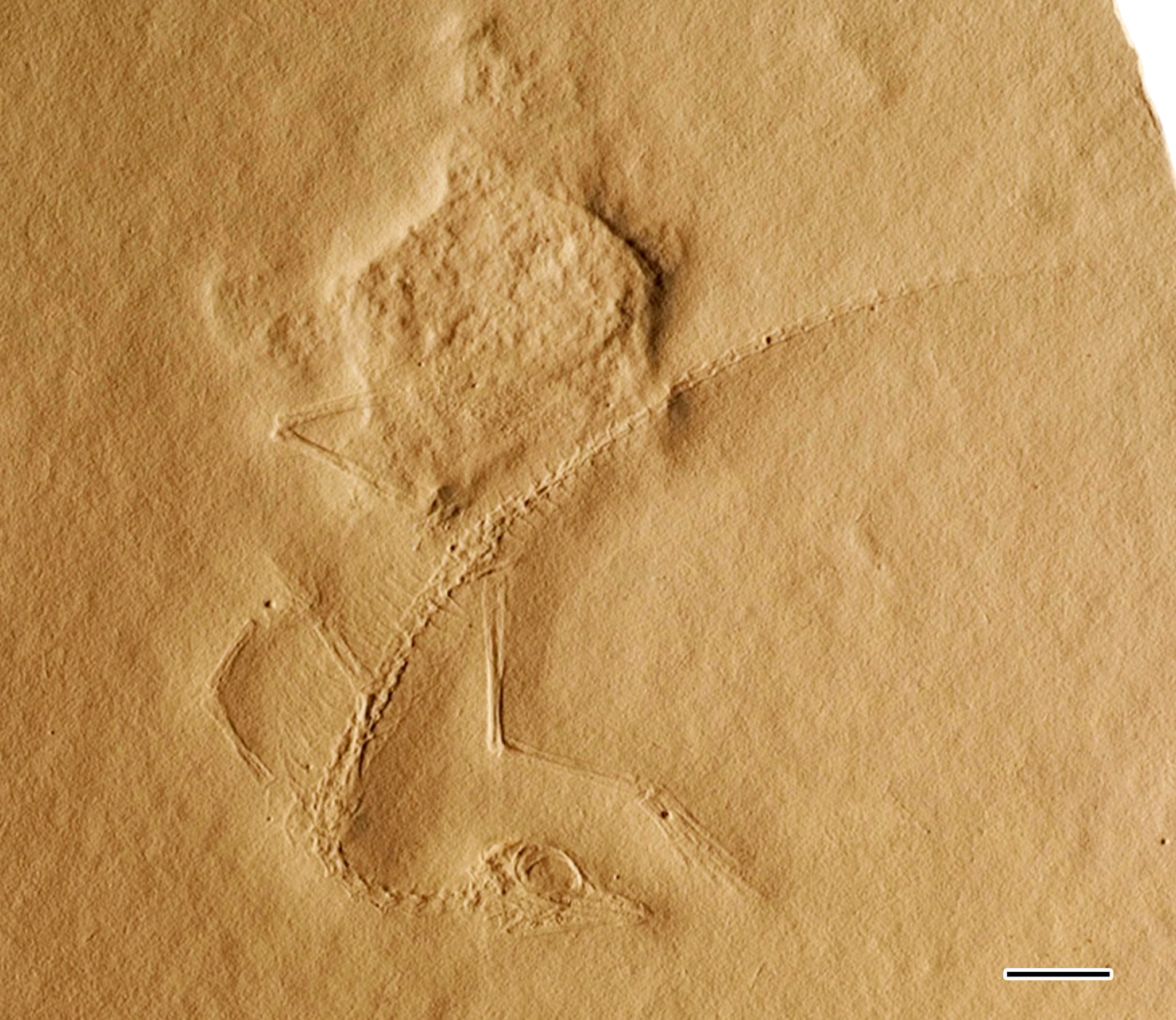
Cosesaurus это род из архозавроморф рептилии вероятно принадлежащий к семье Tanystropheidae.[1] Он известен по ископаемым отпечаткам одного небольшого скелета MGB V1, найденного в Muschelkalk обнажения вблизи муниципалитетов Mont-ral и Альковер в Испания. Эти обнажения датируются Ладинский возраст среднего Триасовый около 242–237 миллионов лет назад. Образец хранится в Museu Martorell (он же Геологический музей Барселоны), который сейчас является частью Museu de Ciències Naturals де Барселона. Плохая сохранность и вероятно малолетний природа образца привела к анатомии Cosesaurus ошибочно идентифицирован несколькими разными источниками. Например, Пол Элленбергер утверждал, что это был предок птицы в 1970-х, в то время как Дэйв Питерс утверждал, что это был птерозавр предка в 2000 году. Оба эти утверждения противоречат господствующим научным теориям о происхождении той или иной группы и других палеонтологи те, кто изучают образец, не могут найти черты, которые, по сообщениям Элленбергера или Петерса, присутствуют.[2][1] Таким образом, рассматриваются гипотезы Элленбергера и Петерса. крайние теории с сомнительной научной обоснованностью из-за их низкого воспроизводимость. Основные гипотезы об отношениях Cosesaurus в целом согласен, что это "проторозавр", особенно танистрофеид, тесно связанный с длинношеими рептилиями, такими как Макрокнемус, Танитрахелос, Танистрофей, или же Лангобардизавр.[3][1][4]
История
Из-за плохой сохранности в зернистости осадочная порода, окаменелость была неверно истолкована многими палеонтологами в прошлом. Первый описатель экземпляра, Пол Элленбергер, ошибочно наблюдал птицеподобные черты, которые не могли быть подтверждены более поздними исследованиями. Он опубликовал несколько анализов этого образца в 1974, 1977 и 1993 годах.[5][6] Он предположил, что это был предок птицы, но от этой гипотезы вскоре отказались другие палеонтологи.
Например, в 1979 г. Пол Олсон кратко обсудил образец во время описания малого танистрофеида Танитрахелос. Он обнаружил трудности с диагностикой этого образца по фотографиям Элленбергера, но заметил, что у него много общего с танистрофеидами, такими как Танитрахелос и Танистрофей а не птицы.[3]
Гипотеза Элленбергера потеряла популярность в конце 20-го века из-за растущего числа свидетельств того, что происхождение птиц среди целурозавр динозавры.[7] За это время проведено наиболее полное изучение Cosesaurus поскольку Элленбергер исполнял Хосе Луис Санс & Ньевес Лопес-Мартинес в 1984 году. Они обнаружили, что почти ни одна из функций, которые Элленбергер использовала для связи Cosesaurus птицам действительно были. Они также изучили его пропорции и пришли к выводу, что это, вероятно, ювенильная особь. На основе этой гипотезы они построили схему скелета, показывающую, что взрослый Cosesaurus может выглядеть, учитывая, как меняются пропорции рептилий в процессе развития. Санс и Лопес-Мартинес обнаружили, что ближайшим родственником Cosesaurus, вероятно, был «пролацертид», такой как Макрокнемус.[2]
В 2000 году исследователь Дэйв Питерс опубликовал вариант гипотезы Элленбергера.[8] Петерс утверждал, что обнаружил в образце особенности, похожие на те, которые, по утверждению Элленбергера, связаны Cosesaurus птицам. Однако Петерс вместо этого утверждал, что эти предполагаемые особенности указывают на то, что Cosesaurus был предком птерозавров, а не птиц. Однако палеонтологи, исследовавшие образец, не смогли найти доказательств, подтверждающих утверждения Питерса, и отметили, что его связь между Cosesaurus и птерозавры полагались на плохо построенный филогенетический анализ.[9] Консенсусное мнение, как это объясняется и поддерживается такими исследованиями, как Несбитт (2011)[10] и Эскурра (2016),[11] заключается в том, что птерозавры не были тесно связаны с «проторозаврами», такими как Cosesaurus, а вместо этого продвинуты архозавры тесно связан с динозавры. В 2009 году Петерс опубликовал продолжение своего исследования 2000 года, в котором использовался алгоритм цифрового отслеживания, чтобы подтвердить его утверждения о таких особенностях, как препубальные кости и кожные лоскуты, похожие на оборки. Однако этот метод также подвергался резкой критике из-за того, что он полагался на фотографии в литературе, а не на личные наблюдения или компьютерная томография свидетельство.[12][9]
А докторская степень Тезис опубликовано в 2016 году Франко Саллером, хотя основное внимание уделяется костно-мышечной системы из Macrocnemus bassanii, также предоставлено новое описание Cosesaurus. Этот тезис, написанный на Итальянский, опровергали некоторые особенности образца, заявленные Элленбергером или Петерсом, такие как наличие грудины или более двух позвонков в крестце. Это также дало дополнительные значения для анатомии и биологии представителей этого рода.[1]
Описание
Примерная длина экземпляра составляет 14 сантиметров (5,5 дюйма), хотя отпечатки кончика хвоста слабые, поэтому эта длина могла быть немного больше. Из-за плохой сохранности образца трудно найти окончательные признаки. Элленбергер сообщил о перепончатых лапах и пух Перо отпечатки, отпечатанные среди отпечатков костей,[6] и Peters (2000) утверждали, что Элленбергер вместо этого наблюдал остатки кожи, такие как патагиум или же актин волокна. Однако оба эти утверждения о мягких тканях сомнительны, поскольку другие палеонтологи, проанализировавшие образец, объясняют, что текстуру поверхности скальной породы возле костей нельзя отличить от остальной части скальной плиты.[2][1]
Череп
Морда средней длины, не такая удлиненная, как у ранних тероподных динозавров. Целофиз, но не такой короткий, как у ранних амниот.[2] Некоторые детали черепа можно идентифицировать с абсолютной уверенностью. Череп и особенно глаза были довольно большими пропорционально, хотя это, вероятно, ювенильные черты. Куполообразная крыша черепа была самой тонкой над глазами, а затем расширялась к затылку. Во рту было множество маленьких заостренных зубов, в том числе шесть или семь длинных заостренных зубов. предчелюстная кость. Несколько зубов в задней части рта были большими, треугольными и похожими на лезвия по сравнению с более тонкими зубами на остальной части челюсти.[1] Хотя отпечаток черепа раздроблен, заметна небольшая выемка перед глазом. Элленбергер (1977) утверждал, что эта впадина пробита носом и представляет собой анторбитальное окно, в то время как Петерс (2000) утверждал, что он представляет собой серию окон. Однако внимательное изучение Саллера (2016) показало, что невозможно определить глубину впадины и определить, какие кости ее окружают.[1] Челюстной сустав имеет некоторые особенности, указывающие на то, что Cosesaurus был ящерица, похожие на современных рептилий, таких как ящерицы, динозавры и крокодилы, вместо более примитивных групп амниот, таких как ареоселидийцы и Вейгельты. Эти особенности включают наличие ретроартикулярного отростка нижней челюсти и квадратная кость который зазубрен сзади и виден снаружи черепа.[13] Однако гибкость квадрата (и, следовательно, потенциал черепной кинезис) неясно.[2]
Позвонки
Шея умеренно длинная, примерно такой же длины, что и череп, и больше половины длины тела. Он образован 7 или 8 удлиненными позвонками, как у многих других архозавроморфов, таких как ранние динозавры, Пролацерта, и Макрокнемус. Подобно другим «проторозаврам», но в отличие от птиц и птерозавров, шейки матки (шейные позвонки) прикреплены к очень длинным и тонким шейные ребра. Хотя шея короткая по сравнению с другими танистрофеидами, Sanz & López-Martínez (1984) и Saller (2016) отмечают, что шеи танистрофеидов имеют тенденцию становиться пропорционально длиннее с возрастом. Поэтому короткая шея Cosesaurus (по стандартам танистрофеидов), вероятно, является ювенильным признаком.[2][1]
Позвонки тела были короче шейных, примерно равной их высоте. Спинные ребра короткие и тонкие. Элленбергер (1977) предположительно обнаружил, что Cosesaurus обладал тремя сросшимися крестцовый (бедренные) позвонки, а Петерс (2000) утверждал, что присутствовали четыре позвонка. Однако оба эти утверждения были опровергнуты Саллером (2016). Хотя он и заметил, что в тазобедренной области есть место для трех крестцовых позвонков. Cosesaurus, он также показал, что присутствовали только две пары крестцовых ребер и что качество сохранности было слишком плохим, чтобы разглядеть швы внутри крестца, чтобы поддержать утверждения Элленбергера или Петерса. Следовательно, Cosesaurus вероятно, имел только два крестцовых позвонка, как и у большинства рептилий.[1]
Хвост длинный, состоит из 40 и более удлиненных позвонков. У первых были тонкие ребра, длина которых постепенно уменьшалась.[1] Элленбергер утверждал, что позвонки срослись в стержневидный пигостиль и отсутствовали гемальные шипы (шевроны), но оба они позже были развенчаны Сансом и Лопес-Мартинесом (1984).[2] и Саллер (2016).
Грудной пояс и передние конечности
Грудной (плечо) пояс - одна из наиболее часто ошибочно идентифицируемых областей Cosesaurus образец. Элленбергер (1977) утверждал, что птицеподобный грудина (нагрудник) и фуркула (поперечный рычаг) присутствовали, но эти особенности не были обнаружены Сансом и Лопес-Мартинесом (1984).[2] Саллер (2016) подробно остановился на ошибочной идентификации Элленбергера. Он объяснил, что «фуркула» была просто парой несращенных ключиц, как у других рептилий, и что «грудина» была просто закругленной нижней частью коракоидный кости. Петерс (2000) также идентифицировал «грудину» Элленбергера как коракоиды, но он ошибся, предполагая, что грудина присутствовала независимо, поскольку Саллер не смог найти никаких доказательств существования кости в Cosesaurus.[1] Олсон (1979) обратил внимание на плечевой пояс танистрофеидов.[3]
Передние конечности намного меньше задних, но все же приличного размера по стандартам большинства рептилий. В плечевая кость (кость плеча) длинная и прямая, с плоскими суставными поверхностями и серединой, которая немного тоньше кончиков кости. Лучевая и локтевая кости (кости предплечья) похожи по строению. Рука очень длинная, с пятью тонкими пальцами. Третий палец самый длинный, и его длина в сочетании с ладонью превышает длину предплечья.[1]
Тазовый пояс и задние конечности
Бедра и тазовый пояс плохо сохранились. В подвздошная кость, возможно, единственная видимая часть таза, необычно удлинена в Cosesaurus.[1] Олсон (1979) заметил, что у основания хвоста могли быть большие изогнутые кости, похожие на "гетеротопный"кости Танитрахелос и Танистрофей. Эти кости могли быть связаны с репродуктивными процессами. Однако Олсон объяснил, что его наблюдения Cosesaurus были облегчены оригинальными зернистыми фотографиями Элленбергера, и что новые наблюдения, сделанные с использованием таких методов, могут быть сомнительными.[3] Хотя Петерс идентифицировал предполагаемые птерозавроподобные «препубические кости» в Cosesaurus, Даррен Нейш не смог найти четких доказательств наличия таких особенностей, используя фотографии Петерса,[9] и Саллер (2016) не упомянули о заявлениях Петерса во время изучения образца.[1]
Задние конечности были длиннее передних конечностей и туловища в целом, особенности, которые, возможно, отразились на биологии животного. Кости ног были длинными и прямыми, похожими по строению на кости рук. В проксимальный часть бедренная кость (бедренная кость) была шире, чем дистальный часть, как и у танистрофеидов. Четыре из пяти плюсневые кости (основные кости стопы) были длинными, прямыми и тесно связанными друг с другом. Они соединялись с длинными пальцами ног с заостренными когти, причем четвертый палец является самым длинным. Пятая (крайняя) плюсневая кость намного короче и шире, приобретает «крючковатую» форму. Согласно Олсону (1979), пятый палец стопы мог иметь удлиненную плюсневую кость. фаланга (кость пальца), чтобы противодействовать сморщенной пятой плюсне.[3] Эта особенность уникальна для большинства танистрофеидов, за исключением Макрокнемус.[11] Подозрения Олсона были позже подтверждены Саллером (2016), изучившим этот образец.[1]
Классификация
В 1977 году Элленбергер предложил Cosesaurus был предком современных птицы.[6] Эта публикация последовала за описанием похожего на птицу теропод, Дейноних, но он появился задолго до теропод происхождение птиц было широко признано. В этой публикации Элленбергер интерпретировал следующие черты окаменелости: лопатка, а фуркула (поперечный рычаг), килевидная грудина, клювовидные челюсти, ретровертированный лобок и хвост перья. Однако любое другое научное исследование, включая Cosesaurus поскольку Элленбергер (1977) обнаружил, что многие из этих признаков были неправильно идентифицированы и что они вряд ли связаны с птицами.
Исследования, проведенные после Элленбергера (1977), интерпретировали Cosesaurus как "пролацертиформный"или" проторозавр ". Эти термины относятся к определенной группе рептилий с удлиненной шеей, такой как Пролацерта, Макрокнемус, и Танистрофей. Эта группа подверглась значительному пересмотру как внутренне (какие таксоны ей принадлежат), так и внешне (где она принадлежит в генеалогическом древе рептилий). До конца 20-го века «пролацертиформы» были перемещены вокруг рептилий, и некоторые источники считали их предками ящериц. Эти источники включают в себя переописателей Cosesaurus, Санс и Лопес-Мартинес (1984).[2] Однако к концу 1980-х годов был достигнут консенсус, который позиционировал «пролацертиформ» как архозавроморфов, что означало, что они были ближе к архозавры такие как крокодилы и динозавры скорее, чем лепидозавры Такие как ящерицы или змей.[14][13]
Однако термин "Prolacertiformes" распался в 1998 году, когда Дэвид Дилкс определил, что Пролацерта на самом деле не был близким родственником других "пролацертиформ", а был более подопечный архозавроморф.[15] Это открытие привело к тому, что специалисты по архозавроморфам отказались от термина «Prolacertiformes», заменив его термином «Protorosauria», который охватывает многих рептилий, которые когда-то считались близкими к Пролацерта. Cosesaurus сохранился среди проторозавров, хотя его положение в группе обычно не проверяется филогенетическим анализом из-за его фрагментарности. Олсон (1979) предположил, что он находится среди танистрофеидов, группы специализированных, возможно, полуводных проторозавров, включая Танитрахелос и Танистрофей.[3] Такую интерпретацию поддержал и Эскурра. и другие. в 2017 году.[4] Санс и Лопес-Мартинес (1984) определили его как «пролацертид», такой как Макрокнемус и Пролацерта, хотя от этого термина в основном отказались по тем же причинам, что и «Prolacertiformes». Макрокнемус теперь также классифицируется как танистрофеид. Танистрофеиды иногда оказываются венцом других «проторозавров», хотя и не в такой степени, как Пролацерта.[11]
Хотя Санс и Лопес-Мартинес (1984) утверждали, что ближайший родственник Cosesaurus был Макрокнемус на основе их пропорционального анализа,[2] Олсон (1979) и Саллер (2016) поместили его в число более продвинутых представителей Tanystropheidae. Они утверждали, что метатарзальная первая фаланга пятого пальца стопы лишает силы связь с Макрокнемус конкретно.[3][1]
В диссертации Саллера (2016) использовались два различных филогенетических анализа предыдущих авторов для изучения взаимоотношений «проторозавров». Первый из этих анализов был проведен Дилксом (1998), исследованием, которое, как известно, устранило монофилию "Prolacertiformes".[15] Однако, когда анализ был изменен для включения большего количества танистрофеидов и проведен Саллером, результаты были довольно неубедительными. Самые экономные деревья Саллера (MPT) не могли прийти к согласию относительно структуры танистрофеидов, более продвинутых, чем Макрокнемус. MPT - это родословные, которые включают наименьшее количество «шагов» (эволюционных переходов, когда черты приобретаются и теряются). Под руководством бритва Оккама, MPT являются наиболее точным описанием эволюционных отношений. Однако шесть MPT Саллера расходились во мнениях относительно структуры Tanystropheidae. Единственный результат, найденный всеми шестью MPT, заключался в том, что Макрокнемус был самым базальным членом семьи.[1]
Стремясь получить более убедительные результаты, Саллер добавил необычный дрепанозавр Мегаланкозавр к анализу в надежде, что это прояснит отношения. Хотя некоторые части анализа были сделаны более убедительными (например, структура Archosauromorpha была близка к найденной Ezcurra [2016][11]) внутренняя структура Protorosauria и Tanystropheidae была не более убедительной, чем когда анализ проводился без Мегаланкозавр.
Эта проблема побудила Саллера использовать второй филогенетический анализ. Этот анализ, первоначально созданный Притчардом и другие. (2015), особое внимание уделялось танистрофеидам.[16] Этот анализ обнаружил три MPT. Хотя структура Archosauromorpha снова оказалась неубедительной, все три MPT соответствовали структуре Tanystropheidae. Одним из самых интересных результатов было то, что Cosesaurus был помещен в качестве сестринского таксона в Лангобардизавр, необычный танистрофеид, который, возможно, был двуногим и травоядный. Об этой связи сообщили три синапоморфии (производные отличительные признаки): шейные и спинные позвонки с плоскими передний суставы, дорсальные кости с низкими нервными шипами и слабо расширенные дистальные мыщелки бедренной кости.[1]
Ниже приведен результат использования метода Притчарда Саллером (2016). и другие. (2015) анализ. Следующее кладограмма представляет собой строгое согласованное дерево, что означает, что это комбинация трех MPT, которая сокращает спорные области до политомия во избежание несогласия с каким-либо конкретным MPT:[1]
| Архозавроморфа |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Палеобиология
Передвижение
Задние лапы Cosesaurus особи длиннее передних конечностей, что указывает на то, что это возможно, но не определенно, двуногий. Санс и Лопес-Мартинес (1984) использовали пропорциональный инструмент, известный как «Индекс Гальтона», чтобы исследовать такую возможность. Создано Питер Гальтон в исследовании 1976 г. прозауроподов (базальные зауроподоморфы), этот индекс сравнивает длину всей задней конечности с длиной тела животного (не включая шею или хвост). В Cosesaurus образец имел отношение 1,21, что было больше, чем только четвероногий эолацертилы (примитивный чешуйчатый родственников), но меньше, чем только двуногие ранние теропод, такие как Целофиз. Это может указывать на некоторую степень двуногого движения, но предполагается, что этот экземпляр является молодым. В современном крокодилыиндекс Гальтона сокращается с 1,06 до 0,79 между птенцами и взрослыми особями. Если соотношение уменьшается до такой же степени в Cosesaurus люди, они, вероятно, вышли из диапазона, позволяющего двуногие, к тому времени, когда они достигнут совершеннолетия.[2]
Рекомендации
- ^ а б c d е ж грамм час я j k л м п о п q р s Саллер, Франко (15 апреля 2016 г.). "Anatomia, paleobiologia e filogenesi di Macrocnemus bassanii Nopcsa 1930 (Reptilia, Protorosauria) " (PDF). Диссертация: 1–206. Дои:10.6092 / unibo / amsdottorato / 7449.
- ^ а б c d е ж грамм час я j k Sanz, J.L .; Лопес-Мартинес, Н. (1984). "Пролацертиды лепидозавров Cosesaurus aviceps Элленбергер и Вильяльта, заявленные «протоавиане» из среднего триаса Испании ». Geobios. 17 (6): 747–755. Дои:10.1016 / S0016-6995 (84) 80119-9.
- ^ а б c d е ж грамм Олсон, Пол Э. (3 мая 1979 г.). «Новый водный эозухий из Ньюаркской супергруппы (поздний триас - ранняя юра) Северной Каролины и Вирджинии» (PDF). Postilla. 176: 1–13.
- ^ а б Эскурра, Мартин; Фортуни, Жозеп; Муджал, Эудальд; Болет, Арнау (2017). «Первые прямые архозавроморфные останки переходного периода между ранним и средним триасом Пиренейского полуострова» (PDF). Palaeontologia Electronica. 20 (3): 1–10. Дои:10.26879/686.
- ^ Элленбергер, Пол; де Вильяльта, Хосе Ф. (1974). "Sur la présence d'un ancètre possible des Oiseaux dans le Muschelkalk supérieur de Catalogne (Espagne). Note preliminaire". Acta Geológica Hispánica. 9 (5): 162–168.
- ^ а б c Элленбергер, П.-П. (1977). "Quelques precision Sur l'anatomie et la place systematique tres speciale de". Cosesaurus aviceps (Ladinien superieur de Montral, Каталогн) ". Caud. Geologica Iberica. 4: 169–188.
- ^ Padian, K .; Чиаппе, Л.М. (1998). «Происхождение и ранняя эволюция птиц». Биологические обзоры. 73 (1): 1–42. Дои:10.1111 / j.1469-185X.1997.tb00024.x.
- ^ Питерс, Дэвид (декабрь 2000 г.). «Повторное исследование четырех пролацертиформ с последствиями для филогенеза птерозавров». Rivista Italiana di Paleontologia e Stratigrafia (Исследования в области палеонтологии и стратиграфии). 106 (3): 293–336. ISSN 2039-4942.
- ^ а б c Нейш, Даррен (3 июля 2012 г.). «Почему мир должен игнорировать ReptileEvolution.com». Сеть блогов Scientific American. Архивировано из оригинал 7 февраля 2017 г.
- ^ Несбитт, С.Дж. (2011). «Ранняя эволюция архозавров: отношения и происхождение основных клад» (PDF). Бюллетень Американского музея естественной истории. 352: 1–292. Дои:10.1206/352.1. HDL:2246/6112.
- ^ а б c d Эскурра, Мартин Д. (28 апреля 2016 г.). «Филогенетические взаимоотношения базальных архозавроморф с упором на систематику архозавриформ протерозухий». PeerJ. 4: e1778. Дои:10.7717 / peerj.1778. ISSN 2167-8359. ЧВК 4860341. PMID 27162705.
- ^ Беннет, С. С. (2005). Наука о птерозаврах или фантазия о птерозаврах? (PDF). Доисторические времена. 70. С. 21–23, 40.
- ^ а б Эванс, Сьюзан Э. (1988). «Ранняя история и отношения Диапсиды». В Бентоне, Майкл Дж. (Ред.). Филогения и классификация четвероногих.. Оксфорд: Clarendon Press. С. 221–260.
- ^ Бентон, Майкл Дж. (Июнь 1985 г.). «Классификация и филогения диапсидных рептилий». Зоологический журнал Линнеевского общества. 84 (2): 97–164. Дои:10.1111 / j.1096-3642.1985.tb01796.x.
- ^ а б Дэвид М. Дилкс (1998). "Раннетриасовый ринкозавр Mesosuchus browni и взаимоотношения базальных рептилий архозавроморф ». Философские труды Лондонского королевского общества, серия B. 353 (1368): 501–541. Дои:10.1098 / рстб.1998.0225. ЧВК 1692244.
- ^ Причард, Адам С.; Тернер, Алан Х .; Nesbitt, Sterling J .; Irmis, Randall B .; Смит, Натан Д. (04.03.2015). «Танистрофеиды позднего триаса (Reptilia, Archosauromorpha) из северного Нью-Мексико (пачка окаменевшего леса, формация Чинл) и биогеография, функциональная морфология и эволюция Tanystropheidae». Журнал палеонтологии позвоночных. 35 (2): e911186. Дои:10.1080/02724634.2014.911186. ISSN 0272-4634.
внешняя ссылка
- Фотография единственного известного экземпляра, опубликованная в твиттер-аккаунте MCNB.
- Более резкое изображение, опубликованное Хосепом Бисбалом










{kind=link}
{kind=link}