WikiDer > Жук
| Жук | |
|---|---|
 | |
| По часовой стрелке сверху слева: самка золотистого оленя (Lamprima aurata), Жук-носорог (Мегасома sp.), длинноносый долгоносик (Гемистический носорог), ковбойский жук (Хондропига спинная), и разновидности Амблитель. | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Членистоногие |
| Учебный класс: | Насекомое |
| (без рейтинга): | Эндоптеригот |
| Заказ: | Жесткокрылые Линней, 1758 |
| Подзаказы | |
Жуки группа насекомые которые образуют порядок Жесткокрылые /koʊляˈɒптərə/, в суперзаказе Эндоптеригот. Их передняя пара крыльев закалена в подкрылки, надкрылья, что отличает их от большинства других насекомых. Жесткокрылые, насчитывающие около 400 000 видов, являются крупнейшими из всех отрядов, составляя почти 40% описанных насекомых и 25% всех известных форм жизни животных; новые виды обнаруживаются часто. Самый большой из всех семьи, то Curculionidae (долгоносики), насчитывающие около 83 000 особей, принадлежат к этому отряду. Встречается почти во всех средах обитания, кроме моря и полярные регионы, они взаимодействуют со своими экосистемы несколькими способами: жуки часто питаются растениями и грибы, расщеплять остатки животных и растений и есть другие беспозвоночные. Некоторые виды являются серьезными сельскохозяйственными вредителями, например, Колорадский жук, а другие, такие как Coccinellidae (божьи коровки или божьи коровки) едят тля, щитовки, трипсы, и другие насекомые-растения, которые повреждают посевы.
Жуки обычно особенно тяжело переносят экзоскелет в том числе надкрылья, хотя некоторые, такие как бродячие жуки имеют очень короткие надкрылья, а волдыри имеют более мягкие надкрылья. Генерал анатомия жука является довольно однородным и типичным для насекомых, хотя есть несколько примеров новизны, таких как адаптации в водяные жуки которые задерживают пузырьки воздуха под надкрыльями для использования во время ныряния. Жуки эндоптериготы, а значит, они проходят полную метаморфоза, с рядом заметных и относительно резких изменений в строении тела между вылуплением и взрослением после относительно неподвижного куколка сцена. Некоторые, например жуки-олени, иметь отмеченный половой диморфизм, самцы, обладающие чрезвычайно увеличенными челюсти которые они используют для борьбы с другими самцами. Многие жуки апосематический, с яркими цветами и узорами, предупреждающими об их токсичности, в то время как другие безвредны Бейтсовская имитация таких насекомых. Многие жуки, в том числе обитающие в песчаных местах, обладают эффективным камуфляж.
Жуки видны в человеческой культуре, от священные скарабеи из древний Египет к жук искусство и использовать как домашние питомцы или же борьба с насекомыми для развлечений и азартных игр. Многие группы жуков имеют яркую и привлекательную окраску, что делает их объектами коллекционирования и декоративного оформления. Более 300 видов используется как еда, в основном как личинки; широко потребляемые виды включают мучные черви и Жук-носорог личинки. Однако основное влияние жуков на жизнь человека проявляется в сельском, лесном и садоводческом хозяйстве. вредители. К серьезным вредителям относятся: долгоносик хлопка, Колорадский жук, то кокосовый жук, а горный сосновый жук. Однако большинство жуков не причиняют экономического ущерба, а многие, например, божьи коровки и навозные жуки полезны, помогая бороться с насекомыми-вредителями.
Этимология

Название таксономического отряда Coleoptera происходит от Греческий колеоптерос (κολεόπτερος), присвоенный группе Аристотель для них надкрылья, закаленные щитковидные передние крылья, из Колеос, оболочка и птерон, крыло. Английское название beetle происходит от Древнеанглийский слово Bitela, немного кусачий, связанный с битан (кусать),[2][3] ведущий к Средний английский Betylle.[4] Другое древнеанглийское название жука - eafor, chafer, используется в таких именах, как хуй, из прото-германского *кебро («жук»; ср. нем. Кафер, Нидерландский язык Кевер).[5]
Распространение и разнообразие
Жуки - безусловно, самый крупный отряд насекомых: около 400 000 видов составляют около 40% всех описанных видов насекомых и около 25% всех животных.[1][6][7][8][9][10] Исследование 2015 года предоставило четыре независимых оценки общего числа видов жуков, давая среднюю оценку около 1,5 миллиона с «удивительно узким диапазоном».[11] охватывающий все четыре оценки от минимум 0,9 до максимум 2,1 миллиона видов жуков. В четырех оценках использовались отношения специфичности хозяина (от 1,5 до 1,9 миллиона), отношения с другими таксонами (от 0,9 до 1,2 миллиона), соотношение растение: жук (от 1,2 до 1,3) и экстраполяции, основанные на размере тела по годам описания (1,7 до 2,1 миллиона).[11][12]
Жуки встречаются почти во всех средах обитания, включая пресноводные и прибрежные, везде, где встречается растительная листва, от деревьев и их коры до цветов, листьев и под землей возле корней - даже внутри растений в галлах, в каждой растительной ткани, включая мертвые или разлагающиеся. ед.[13] В пологах тропических лесов обитает большая и разнообразная фауна жуков,[14] включая Carabidae,[15] Chrysomelidae,[16] и Scarabaeidae.[17]
Самый тяжелый жук, на самом деле самая тяжелая стадия насекомого - это личинка из жук-голиаф, Голиаф голиатус, который может достигать массы не менее 115 г (4,1 унции) и длины 11,5 см (4,5 дюйма). Взрослые самцы жуков-голиафов - самые тяжелые жуки во взрослой стадии, их вес составляет 70–100 г (2,5–3,5 унции), а размер достигает 11 см (4,3 дюйма).[18] Взрослый жуки-слоны, Слоны мегасомы и Megasoma actaeon часто достигают 50 г (1,8 унции) и 10 см (3,9 дюйма).[19]
Самый длинный жук - это Жук-геркулес Династия геркулеса, с максимальной общей длиной не менее 16,7 см (6,6 дюйма), включая очень длинные пронотальный Рог. Самый маленький зарегистрированный жук и самое маленькое свободноживущее насекомое (по состоянию на 2015 г.[Обновить]), это перкокрылый жук Scydosella musawasensis который может иметь размер всего 325мкм в длину.[20]
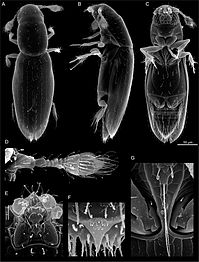
Scydosella musawasensis, самый маленький из известных жуков: масштабная линейка (справа) составляет 50 мкм.

Жук-геркулес, Dynastes hercules ecuatorianus, самый длинный из жуков



Эволюция
Поздний палеозой

Самое древнее из известных ископаемых насекомых, однозначно напоминающих жесткокрылых, происходит из Нижняя пермь о 270 миллион лет назад (mya), хотя эти члены семьи Tshekardocoleidae имеют 13-сегментный усики, надкрылья с более развитым жилкованием и более неравномерной продольной ребристостью, брюшко и яйцеклад заходит за вершину надкрылий. в Пермско-триасовое вымирание в конце перми вымерло около 30% всех видов насекомых, поэтому в летописи окаменелостей насекомых есть только жуки из Нижний триас 220 моя. Примерно в это же время, в позднем триасе, грибок такие виды, как Cupedidae появляются в летописи окаменелостей. В этапах верхнего триаса, кормление водорослями насекомые, такие как Triaplidae и Гидрофилиды начинают появляться вместе с хищными водными жуками. Первые долгоносики, в том числе Obrienidae, появляются рядом с первыми бродячими жуками (Staphylinidae), которые очень напоминают современные виды.[21] Некоторые энтомологи скептически относятся к тому, что такие ранние насекомые так тесно связаны с современными видами, утверждая, что это крайне маловероятно; например, строение метэпистерна предполагает, что Obrienidae могли быть Archostemata, а вовсе не долгоносиками, несмотря на окаменелости с мордами, напоминающими долгоносиков.[22]
В 2009 г. был описан ископаемый жук из Пенсильванский из Mazon Creek, Штат Иллинойс, отодвигая происхождение жуков на более раннюю дату, 318 по 299 моя.[23] Окаменелости того времени были найдены в Азии и Европе, например, в красных сланцевых окаменелостях Нидермошеля недалеко от Майнца, Германия.[24] Другие окаменелости были обнаружены в Оборе, Чехия, и Чекарде на Урале, Россия.[25] Однако есть всего несколько окаменелостей из Северной Америки до средняя пермь, хотя и Азия, и Северная Америка были объединены Euramerica. Первые открытия из Северной Америки, сделанные в формации Веллингтон в Оклахоме, были опубликованы в 2005 и 2008 годах.[21][26]
Как следствие Пермско-триасовое вымираниелетопись окаменелостей насекомых скудна, включая жуков из нижнего триаса.[27] Однако есть несколько исключений, например, в Восточной Европе. На стоянке Бабий Камень в Кузнецкой котловине обнаружены многочисленные окаменелости жуков, в том числе целые экземпляры инфраотряда. Архостемы (например, Ademosynidae, Schizocoleidae), Адефага (например, Triaplidae, Trachypachidae) и Полифага (например, Hydrophilidae, Byrrhidae, Elateroidea).[28] Однако виды из семейств Cupedidae и Шизофороиды отсутствуют на этом участке, тогда как они преобладают на других участках ископаемых из Нижний триас такие как Хей-Яга, Россия, в бассейне Коротаихи.[21]
Юрский
Вовремя Юрский (210 к 145 моя) резко увеличилось разнообразие семейств жуков,[21] включая развитие и рост хищных и травоядных видов. В Хризомелоидея диверсифицировались примерно в то же время, питаясь широким спектром растений-хозяев из саговники и хвойные породы к покрытосеменные.[29] Ближе к верхней юре численность Cupedidae уменьшилась, но увеличилось разнообразие ранних видов, питающихся растениями. Самые современные жуки-растительноядные питаются цветущими растениями или покрытосеменными, чей успех способствовал удвоению числа растительноядных видов в период Средняя юра. Однако увеличение количества семейств жуков в меловом периоде не коррелирует с увеличением количества видов покрытосеменных.[30] Примерно в то же время многочисленные примитивные долгоносики (например, Curculionoidea) и жуков-щелкунов (например, Elateroidea) появившийся. Первые жуки-драгоценности (например, Buprestidae) присутствуют, но они оставались редкими до мелового периода.[31][32][33] Первые жуки-скарабеи не были копрофагами, но предположительно питались гниющей древесиной с помощью грибка; они являются ранним примером мутуалистический отношение.
Есть более 150 важных памятников юрского периода, большинство из которых находятся в Восточной Европе и Северной Азии. Выдающиеся сайты включают Solnhofen в Верхнем Бавария, Германия,[34] Каратау на юге Казахстан,[35] формация иссянь в Ляонин, Северный Китай,[36] а также формация Цзюлуншань и другие ископаемые останки в Монголия. В Северной Америке есть лишь несколько мест с ископаемыми останками насекомых юрского периода, а именно залежи ракушечника в бассейне Хартфорд, бассейн Дирфилд и бассейн Ньюарк.[21][37]
Меловой
В Меловой увидел фрагментацию южной части суши с открытием южной части Атлантического океана и изоляцией Новой Зеландии, в то время как Южная Америка, Антарктида и Австралия стали более отдаленными.[29] Разнообразие Cupedidae и Архостемы значительно уменьшилось. Хищные жужелицы (Carabidae) и стафилиниды (Staphylinidae) начали распределяться по разным образцам; то Carabidae преобладали в теплых регионах, а Staphylinidae и жуки-щелкуны (Elateridae) предпочитали умеренный климат. Точно так же хищные виды Клероида и Cucujoidea охотились на свою добычу под корой деревьев вместе с драгоценные жуки (Buprestidae). Разнообразие жемчужных жуков быстро увеличивалось, поскольку они были основными потребителями древесины,[38] пока усачьи жуки (Cerambycidae) были довольно редки: их разнообразие увеличилось только к концу верхнего мела.[21] Первые жуки-копрофаги происходят из верхнего мела.[39] и, возможно, жили на экскрементах травоядных динозавров.[40] Встречаются первые виды, у которых и личинки, и взрослые особи адаптированы к водному образу жизни. Жуки-вихри (Gyrinidae) были умеренно разнообразными, хотя другие ранние жуки (например, Dytiscidae) были менее разнообразными, причем наиболее распространенными были виды Coptoclavidae, которые охотились на личинок водных мух.[21]Обзор палеоэкологической интерпретации ископаемых жуков из мелового янтаря в 2020 году показал, что сапроксиличность была наиболее распространенной стратегией кормления с грибоядный вид, в частности, кажется, чтобы доминировать.[41]
Многие ископаемые останки по всему миру содержат жуков мелового периода. Большинство из них находятся в Европе и Азии и относятся к зоне умеренного климата в меловом периоде.[36] Участки нижнего мела включают пласты окаменелостей Крато в бассейне Арарипе в Сеара, Северная Бразилия, а также вышележащая формация Сантана; последний в то время находился около экватора. В Испании важные объекты находятся рядом Montsec и Лас Хойас. В Австралии ископаемые пласты Кунварра группы Корумбурра, South GippslandВиктория, заслуживают внимания. Основные участки верхнего мела включают: Кзыл-Джар в Южном Казахстане и Аркагала в России.[21]
Кайнозойский

Окаменелости жуков многочисленны в кайнозое; посредством Четвертичный (до 1,6 млн лет назад) ископаемые виды идентичны живым, а из Поздний миоцен (5,7 млн лет назад) окаменелости все еще настолько близки к современным формам, что, скорее всего, являются предками живых видов. В большие колебания климата Во время четвертичного периода жуки так сильно изменили свое географическое распространение, что нынешнее местонахождение мало что может сказать о биогеографической истории вида. Очевидно, что географическая изоляция популяций часто должна была нарушаться, поскольку насекомые перемещались под влиянием меняющегося климата, вызывая смешение генофондов, быструю эволюцию и вымирание, особенно в средних широтах.[43]
Филогения
Очень большое количество видов жуков создает особые проблемы для классификация. Некоторые семейства содержат десятки тысяч видов, и их необходимо разделить на подсемейства и племена. Это огромное количество привело биолога-эволюциониста Дж. Б. С. Холдейн язвительно, когда некоторые теологи спросили его, что можно сделать вывод о разуме Творца из произведений Его творения, «чрезмерной любви к жукам».[44]Полифага самый крупный подотряд, содержащий более 300 000 описанных видов более чем 170 семейств, в том числе бродячие жуки (Staphylinidae), жуки-скарабеи (Scarabaeidae), волдыри (Meloidae), жуки-олени (Lucanidae) и настоящие долгоносики (Curculionidae).[9][45] Эти группы жуков-полифагов можно определить по наличию шейных склериты (затвердевшие части головы, используемые как точки прикрепления мышц) отсутствуют в других подотрядах.[46]Адефага содержит около 10 семейств преимущественно хищных жуков, включая жужелицы (Carabidae), водяные жуки (Dytiscidae) и жуки-вихри (Gyrinidae). У этих насекомых яички трубчатые и первая грудина брюшной полости (пластинка экзоскелет) делится на заднюю часть тазики (базальные суставы ног жука).[47]Архостемы содержит четыре семейства преимущественно древесных жуков, в том числе сетчатые жуки (Cupedidae) и телефонный жук.[48]У Archostemata есть открытая пластина, называемая метатрочантин, перед базальным сегментом или тазиком задней ноги.[49] Миксофага содержит около 65 описанных видов четырех семейств, в основном очень мелких, в том числе Hydroscaphidae и род Sphaerius.[50] Жуки-миксофаганы маленькие и в основном питаются водорослями. Для их ротового аппарата характерно отсутствие зубцов и подвижный зуб на левой нижней челюсти.[51]
Консистенция жука морфология, в частности наличие у них надкрылий, давно предполагают, что жесткокрылые монофилетический, хотя были сомнения в расположении подзаказы, а именно Адефага, Архостемы, Миксофага и Полифага внутри этого клады.[52][29][53][54][55] Паразиты с перекрученными крыльями, Strepsiptera, считаются сестринской группой жуков, отделившись от них в Ранняя пермь.[54][56][57][58]
Молекулярно-филогенетический анализ подтверждает, что жесткокрылые монофилетики. Duane McKenna et al. (2015) использовали восемь ядерных генов для 367 видов из 172 из 183 семейств жесткокрылых. Они разделили Adephaga на 2 клады, Hydradephaga и Geadephaga, разделили Cucujoidea на 3 клады и поместили Lymexyloidea в Tenebrionoidea. Полифаги датируются триасом. Большинство существующих семейств жуков, по-видимому, возникли в меловом периоде.[58] В кладограмма основан на McKenna (2015).[58] Количество видов в каждой группе (в основном надсемейства) показано в скобках и жирным шрифтом, если оно превышает 10 000.[59] Там, где возможно, приводятся общеупотребительные английские имена Даты происхождения основных групп показаны курсивом в миллионы лет назад (млн лет назад).[59]
| Жесткокрылые |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 285мя |





Внешняя морфология

Жуки обычно отличаются особенно тяжелым экзоскелет и жесткие передние крылья (надкрылья) не годится для полета. Почти у всех жуков челюсти движутся в горизонтальной плоскости. Ротовой аппарат редко бывает присосным, хотя иногда и уменьшен; на максиллах всегда есть щупики. Усики обычно имеют 11 или меньше сегментов, за исключением некоторых групп, таких как Cerambycidae (усики) и Rhipiceridae (жуки-цикады). Тазики ног обычно расположены углубленно в полости тазика. Генитальные структуры у всех современных жуков выдвинуты в последний сегмент брюшка. Личинки жуков часто можно спутать с личинками других групп эндоптериго.[49] Экзоскелет жука состоит из множества пластин, называемых склериты, разделенных тонкими швами. Такая конструкция обеспечивает бронированную защиту при сохранении гибкости. Генерал анатомия жука довольно однородный, хотя специфические органы и придатки сильно различаются по внешнему виду и функциям между многими семьями в порядке. Как и у всех насекомых, тело жуков разделено на три части: голову, грудную клетку и брюшко.[7] Поскольку существует так много видов, идентификация довольно сложна и зависит от атрибутов, включая форму усиков, тарзальные формулы[а] и формы этих маленьких сегментов на ногах, ротовом аппарате и брюшных пластинах (грудины, плевре, тазиках). У многих видов точная идентификация может быть сделана только путем изучения уникальных мужских половых органов.[60]
Голова
.jpg)
Голова с выступающим вперед или иногда опущенным ротовым аппаратом обычно сильно склеротизированный а иногда бывает очень большим.[6] Глаза сложный и может проявлять замечательную приспособляемость, как в случае водных жуков-вертушек (Gyrinidae), где они разделены, чтобы обеспечить обзор как выше, так и ниже ватерлинии. Немного Жуки-усачьи (Cerambycidae) и долгоносиков, а также некоторых светлячков (Rhagophthalmidae) [61] разделили глаза, в то время как у многих глаза с зазубринами, а у некоторых глазки, маленький, простой глаза обычно дальше на затылке (на вершина); они чаще встречаются у личинок, чем у взрослых.[62] Анатомическая организация сложных глаз может быть изменена и зависит от того, является ли вид преимущественно сумеречным, дневным или ночным активным.[63] Глазки встречаются у взрослых ковровых жуков (Dermestidae), некоторые бродячие жуки (Omaliinae), а Derodontidae.[62]

Жук усики являются в первую очередь органами сенсорного восприятия и могут обнаруживать движение, запах и химические вещества,[64] но также может использоваться для физического ощущения среды обитания жука. Семейства жуков могут использовать усики по-разному. Например, при быстром движении тигровые жуки могут плохо видеть и вместо этого жестко держат свои усики перед собой, чтобы избежать препятствий.[65]Некоторые Cerambycidae используют усики для баланса, а волдыри могут использовать их для хватания. Некоторые виды водных жуков могут использовать антенны для сбора воздуха и пропускания его под телом, когда они находятся под водой. Точно так же некоторые семьи используют антенны во время спаривания, а некоторые виды используют их для защиты. В церамбициде Onychocerus albitarsis, антенны имеют структуры, вводящие яд, используемые в защите, что является уникальным среди членистоногие.[66] Антенны сильно различаются по форме, иногда у представителей разных полов, но часто они похожи в пределах одной семьи. Усики могут быть забитый, нитевидный, угловой, в форме бус, гребенчатый (с одной или обеих сторон, двояковыпуклые), или зубчатый. Физические изменения усиков важны для идентификации многих групп жуков. У Curculionidae есть локтевые или коленчатые усики. Перышкообразные усики - это ограниченная форма, встречающаяся у Rhipiceridae и некоторых других семейств. У Silphidae головчатые антенны со сферической головкой на конце. У Scarabaeidae обычно есть пластинчатые усики с конечными сегментами, вытянутыми в длинные плоские структуры, сложенные вместе. У Carabidae обычно есть нитевидные усики. Усики возникают между глазом и нижними челюстями, а у Tenebrionidae они поднимаются перед выемкой, которая нарушает обычно круглый контур сложного глаза. Они сегментированы и обычно состоят из 11 частей, первая часть называется скапусом, а вторая - ножкой. Остальные сегменты вместе называются жгутиком.[64][67][68]
Жуки имеют ротовые органы как у кузнечики. В челюсти выглядят как большие клешни на передней части некоторых жуков. Челюсти представляют собой пару твердых, часто зубчатых структур, которые перемещаются горизонтально, чтобы захватывать, раздавливать или разрезать еду или врагов (см. защита, ниже). Две пары пальцевидных придатков, верхнечелюстные и губные щупики, находятся вокруг рта у большинства жуков и служат для перемещения пищи в рот. У многих видов нижние челюсти имеют половой диморфизм, при этом у самцов они значительно увеличены по сравнению с таковыми у самок того же вида.[6]
Грудная клетка
Грудная клетка сегментированный на две заметные части, про- и птероторакс. Птероторакс - это сросшийся мезо- и заднегрудь, которые обычно разделяются у других видов насекомых, хотя и гибко соединяются с переднегрудием. Если смотреть снизу, то грудная клетка - это та часть, из которой возникают все три пары ног и обе пары крыльев. Живот - это все кзади от грудной клетки.[7] Если смотреть сверху, кажется, что у большинства жуков есть три четких участка, но это обманчиво: на верхней поверхности жука средняя часть представляет собой твердую пластину, называемую переднеспинка, который представляет собой только переднюю часть грудной клетки; задняя часть грудной клетки закрыта жуком крылья. Дальнейшая сегментация обычно лучше всего видна на животе.[69]

Ноги
Многосегментный ноги заканчиваются двумя-пятью небольшими сегментами, называемыми лапками. Как и у многих других отрядов насекомых, у жуков есть когти, обычно одна пара, на конце последнего членика предплюсны каждой ноги. В то время как большинство жуков используют свои ноги для ходьбы, ноги были по-разному приспособлены для других целей. Водные жуки, включая Dytiscidae (ныряющие жуки), Haliplidae, и многие виды Гидрофилиды, ноги, часто последняя пара, приспособлены для плавания, обычно с рядами длинных волос. У самцов ныряющих жуков есть присоски на передних лапах, которые они используют для захвата самок.[70] У других жуков есть роющий ноги расширены и часто колючие при копании. Виды с такими приспособлениями встречаются среди скарабеев, жужелиц и жуки-клоуны (Histeridae). Задние лапы некоторых жуков, например блошиные жуки (у Chrysomelidae) и долгоносиков-блох (в Curculionidae) увеличены бедренные кости, которые помогают им прыгать.[71]
Крылья
.jpg)
Передние крылья жуков не используются для полет, но образуют надкрылья, которые покрывают заднюю часть тела и защищают задние крылья. Надкрылья обычно представляют собой твердые, похожие на раковину структуры, которые необходимо приподнять, чтобы задние крылья могли двигаться для полета.[72] Однако у жуков-солдатиков (Cantharidae) надкрылья мягкие, благодаря чему это семейство прозвали кожекрылыми.[73] К другим мягкокрылым жукам относятся сеткокрылый жук Калоптерон Dispans, у которого есть хрупкие крылья, которые легко ломаются, выделяя химические вещества для защиты.[74]
Летающие крылья жуков пересекаются жилками и складываются после приземления, часто вдоль этих жилок, и хранятся ниже надкрылий. Сгиб (jugum) перепонки у основания каждого крыла.[72] Некоторые жуки утратили способность летать. К ним относятся некоторые жужелицы (Carabidae) и некоторые настоящие долгоносики (Curculionidae), а также обитающие в пустынях и пещерах виды других семейств. У многих два надкрылья слиты вместе, образуя прочный щит над брюшком. В некоторых семьях утрачены и способность летать, и надкрылья, как у светлячки (Phengodidae), где самки похожи на личинок на протяжении всей своей жизни.[75] Наличие надкрылий и крыльев не всегда говорит о том, что жук будет летать. Например, пижма ходит между местами обитания, несмотря на то, что физически способен летать.[76]
Брюшная полость
В брюшная полость это часть позади заднегруди, состоящая из серии колец, каждое из которых имеет отверстие для дыхания и дыхания, называемое дыхальце, составляющие три различных сегментированных склерита: тергум, плевру и грудину. Тергум почти у всех видов перепончатый или обычно мягкий и скрыт крыльями и надкрыльями, когда он не в полете. У некоторых видов плевры обычно маленькие или скрытые, причем каждый плеврон имеет одно дыхальце. Грудина - наиболее видимая часть живота, представляющая собой более или менее склеротизированный сегмент. Сама брюшная полость не имеет придатков, но некоторые (например, Морделлиды) имеют сочленяющиеся доли грудины.[77]
Анатомия и психология

Пищеварительная система
В пищеварительная система жуков в первую очередь адаптирован для растительноядного питания. Пищеварение происходит в основном в переднем отделе. средняя кишка, хотя в хищных группах вроде Carabidae, большая часть пищеварения происходит с помощью ферментов средней кишки. в Elateridae, личинки представляют собой жидкие кормушки, которые экстраорально переваривают пищу, выделяя ферменты.[7] Пищеварительный канал в основном состоит из короткого узкого глотка, широкая экспансия, урожай и слабо развитая желудок. Далее следует средняя кишка, размеры которой у разных видов различаются, с большим количеством слепая кишкаи задняя кишка разной длины. Обычно бывает от четырех до шести Мальпигиевы канальцы.[6]
Нервная система
В нервная система in beetles содержит все типы, встречающиеся у насекомых, в зависимости от вида, от трех грудных и семи или восьми брюшных ганглиев, которые можно различить, до ганглиев, в которых все грудные и брюшные ганглии слиты, образуя сложную структуру.[7]
Дыхательная система

Как и большинство насекомых, жуки вдыхают воздух, так как кислород он содержит, и выдох углекислый газ, через трахеальная система. Воздух попадает в организм через дыхальца, и циркулирует в гемоцеле в системе трахеи и трахеолы, через стенки которых могут диффундировать газы.[7]
Жуки-ныряльщики, такие как Dytiscidae, когда ныряют, носите с собой пузырь воздуха. Такой пузырь может удерживаться под надкрыльями или напротив тела специальными средствами. гидрофобный волосы. Пузырек покрывает, по крайней мере, некоторые дыхальца, позволяя воздуху проникать в трахеи.[7] Функция пузыря - не только удерживать воздух, но и действовать как физическая жабра. Воздух, который он улавливает, находится в контакте с насыщенной кислородом водой, поэтому, поскольку потребление животного истощает кислород в пузыре, больше кислорода может диффундировать, чтобы пополнить его.[78] Углекислый газ более растворим в воде, чем кислород или азот, поэтому он легко диффундирует быстрее, чем в воде. Азот - самый распространенный газ в пузырьке и наименее растворимый, поэтому он составляет относительно статический компонент пузырька и действует как стабильная среда для скопления и прохождения дыхательных газов. Жуку достаточно случайных выходов на поверхность, чтобы восстановить строение пузыря.[79]
Сердечно-сосудистая система
Как и другие насекомые, у жуков есть открытые системы кровообращения, на основе гемолимфа а не кровь. Как и у других насекомых, к дорсальной стенке ствола прикреплено сегментированное трубчатое сердце. гемоцель. Имеет парные входы или Остия через определенные промежутки времени по ее длине, и гемолимфа циркулирует из основной полости гемоцеля и выходит через переднюю полость головы.[80]
Специализированные органы
Разные железы специализируются на разных феромонах, привлекающих партнеров. Феромоны из видов Rutelinae производятся из эпителиальные клетки выстилает внутреннюю поверхность апикальных сегментов брюшка; феромоны на основе аминокислот Melolonthinae образуются из выворачивающихся желез на верхушке брюшной полости. Другие виды производят разные типы феромонов. Дерместиды производить сложные эфиры, и виды Elateridae производить альдегиды, производные жирных кислот и ацетаты.[7] Чтобы привлечь партнера, светлячки (Lampyridae) используют модифицированные клетки жирового тела с прозрачными поверхностями, покрытыми отражающими кристаллами мочевой кислоты, чтобы производить свет путем биолюминесценция. Легкое производство высокоэффективно за счет окисления люциферин катализируется ферментами (люциферазы) в присутствии аденозинтрифосфат (АТФ) и кислород, производя оксилюциферин, углекислый газ и свет.[7]
Барабанные органы или органы слуха, состоящие из мембраны (барабанной перепонки), натянутой на каркас, поддерживаемой воздушным мешком и связанными с ней сенсорными нейронами, встречаются в двух семьях.[81] Несколько видов рода Cicindela (Carabidae) имеют органы слуха на дорсальных поверхностях их первых брюшных сегментов под крыльями; два племени в Dynastinae (в рамках Scarabaeidae) имеют органы слуха прямо под щитками переднеспинки или перепонками шеи. Обе семьи чувствительны к ультразвуковым частотам, и есть веские доказательства, указывающие на то, что они функционируют, чтобы обнаруживать присутствие летучих мышей с помощью их ультразвуковой эхолокации.[7]
Размножение и развитие
Жуки являются членами суперзаказ Эндоптеригот, и соответственно большинство из них проходят полную метаморфоза. Типичная форма метаморфоза у жуков проходит четыре основных этапа: яйцо, то личинка, то куколка, а имаго или взрослый. Личинок обычно называют личинки куколку иногда называют куколкой. У некоторых видов куколка может быть заключена в кокон, созданный личинкой ближе к концу своего последнего периода жизни. возраст. Некоторые жуки, например, типичные представители семейства Meloidae и Рипифориды, идите дальше, проходя гиперметаморфоз в котором первая возрастная стадия принимает форму триунгулин.[82]
Спаривание

У некоторых жуков сложное брачное поведение. Феромон Общение часто важно при поиске партнера. Разные виды используют разные феромоны. Жуки-скарабеи, такие как Rutelinae использовать феромоны, полученные из синтез жирных кислот, в то время как другие скарабеи, такие как Melolonthinae использовать аминокислоты и терпеноиды. Еще один способ, которым жуки находят себе пару, можно увидеть в светлячки (Lampyridae), которые биолюминесцентный, с брюшными светообразующими органами. Самцы и самки вступают в сложный диалог перед спариванием; Каждый вид имеет уникальное сочетание схемы полета, продолжительности, состава и интенсивности излучаемого света.[7]
Перед спариванием самцы и самки могут стричь или вибрировать предметы, на которых они находятся. У Meloidae самец забирается на спину самки и поглаживает усиками ее голову, щупальца и усики. В Eupompha, самец проводит усиками вдоль продольной вершины. Они могут вообще не спариваться, если не выполняют прекопуляционный ритуал.[7] Такое брачное поведение может быть различным среди рассредоточенных популяций одного и того же вида. Например, вязка русский население пижма (Chysolina graminis) предшествует тщательно продуманный ритуал, в котором самец постукивает своими антеннами по глазам, переднеспинке и усикам самки, что не проявляется в популяции этого вида в объединенное Королевство.[83]
Конкуренция может играть роль в брачных ритуалах таких видов, как жуки-закопатели (Никрофор), насекомые сражаются, чтобы определить, какие из них могут спариваться. Многие жуки-самцы территориальный и яростно защищают свои территории от вторжения мужчин. У таких видов самец часто имеет рога на голове или грудной клетке, поэтому длина его тела больше, чем у самки. Спаривание обычно быстрое, но в некоторых случаях длится несколько часов. Во время совокупления сперматозоиды передаются женщине в удобрять яйцо.[6]
Жизненный цикл

Яйцо
Практически все жуки откладывают яйца, хотя некоторые мирмекофил Aleocharinae и немного Chrysomelinae которые живут в горах или в субарктике, яйцекладущиеоткладывают яйца, которые вылупляются почти сразу. Яйца жуков обычно имеют гладкую поверхность и мягкие, хотя Cupedidae есть твердые яйца. Яйца у разных видов сильно различаются: яйца, как правило, маленькие у видов со многими возрастами (личиночные стадии) и у тех, которые откладывают большое количество яиц. Самка может отложить от нескольких десятков до нескольких тысяч яиц в течение своей жизни, в зависимости от степень родительской заботы. Это варьируется от простой откладки яиц под листом до родительской заботы, обеспечиваемой жуки-скарабеи, которые содержат, кормят и защищают своих детенышей. В Attelabidae скручивайте листья и кладите яйца внутрь рулета для защиты.[7][84]
Личинка
В личинка обычно основной этап питания жука жизненный цикл. Личинки, как правило, обильно питаются после выхода из яиц. Некоторые питаются внешними растениями, например, некоторыми листоедами, в то время как другие питаются из своих источников пищи. Примеры внутренних питателей наиболее Buprestidae и усачьи жуки. Личинки многих семейств жуков хищны, как и взрослые особи (жужелицы, божьи коровки, стафилококки). Личиночный период варьируется между видами, но может достигать нескольких лет. Личинки кожные жуки при голодании претерпевают определенную степень обратного развития, а затем возвращаются к ранее достигнутому уровню зрелости. Цикл можно повторять много раз (см. Биологическое бессмертие).[85] Морфология личинок сильно различается у разных видов, с хорошо развитыми и склеротизированными головами, различимыми грудными и брюшными сегментами (обычно десятым, но иногда восьмым или девятым).[6]
.jpg)
Личинки жуков можно отличить от личинок других насекомых по их затвердевшим, часто затемненным головам, наличию жевательных частей рта и дыхальца по бокам их тел. Как и взрослые жуки, личинки различаются по внешнему виду, особенно между семействами жуков. Жуки с несколько приплюснутыми, очень подвижными личинками включают жужелицы и бродячие жуки; их личинки описываются как камподевидные. Некоторые личинки жуков напоминают окаменевших червей с темными головными капсулами и крошечными ногами. Это личинки элатериформной формы, встречающиеся в жук-щелкун (Elateridae) и чернотелка (Tenebrionidae) семей. Некоторые личинки жуков-щелкунов элатериформной формы известны как проволочники. Жуки в Scarabaeoidea имеют короткие толстые личинки, описываемые как скарабеи, более известные как личинки.[86]
Все личинки жуков проходят несколько возрастов, которые являются этапами развития между каждым линька. У многих видов личинки просто увеличиваются в размерах с каждым последующим возрастом по мере того, как потребляется больше пищи. Однако в некоторых случаях происходят более резкие изменения. Среди определенных семейств или родов жуков, особенно тех, которые демонстрируют паразитический образ жизни, первый возраст ( планидиум) очень мобильна для поиска хозяина, в то время как следующие возрастные группы более малоподвижны и остаются на своем хозяине или внутри него. Это известно как гиперметаморфоз; это происходит в Meloidae, Micromalthidae, и Ripiphoridae.[87] Жук пузыря Epicauta vittata (Meloidae), например, имеет три различных личиночных стадии. Его первый этап, триунгулин, имеет более длинные ноги, чтобы отправиться на поиски яиц кузнечиков. После кормления в течение недели он линяет до второй стадии, называемой карабовидной стадией, которая напоминает личинку особи. жук-жук. Еще через неделю он линяет и принимает вид личинка скарабеи - стадия скарабеевидов. Предпоследняя личиночная стадия - это псевдокуколка или грубоватая личинка, которая перезимует и окуклится до следующей весны.[88]
Личиночный период может широко варьироваться. Грибок, питающий стафилиниды Phanerota fasciata претерпевает три линьки за 3,2 дня при комнатной температуре, в то время как Анизотома sp. (Leiodidae) завершает личиночную стадию в плодовом теле слизистой плесени за 2 дня и, возможно, представляет собой наиболее быстрорастущих жуков. Жуки-дерместиды, Trogoderma inclusum могут оставаться в расширенном личиночном состоянии в неблагоприятных условиях, даже уменьшая свои размеры между линьками. Сообщается, что личинка выжила в закрытом контейнере 3,5 года.[7]
Куколка и взрослый

Как и все эндоптериготы, личинки жуков окукливаются, и от них куколки появляются полностью сформировавшиеся половозрелые взрослые жуки или имаго. Куколки никогда не имеют нижних челюстей (они адектики). У большинства куколок придатки не прикреплены к телу и, как говорят, исключать; у некоторых жуков (Staphylinidae, Ptiliidae и т. д.) отростки срослись с телом (называемые обнаруживать куколки).[6]
Продолжительность жизни взрослых особей очень разная, от недель до лет, в зависимости от вида.[6][49] Некоторые жуки-древоточцы могут иметь очень долгий жизненный цикл. Считается, что когда мебель или древесина заражаются личинками жуков, древесина уже содержала личинки при первой распиловке. А береза книжный шкаф 40 лет выпущен взрослый Eburia quadrigeminata (Cerambycidae), пока Buprestis aurulenta и другие Buprestidae задокументированы как возникшие спустя 51 год после изготовления деревянных изделий.[89]
Поведение
Передвижение

Надкрылья позволяют жукам как летать, так и перемещаться в ограниченном пространстве, делая это, складывая нежные крылья под надкрыльями, когда они не летают, и складывая крылья непосредственно перед взлетом. Раскладывание и складывание крыльев осуществляется мышцами, прикрепленными к основанию крыла; пока сохраняется напряжение на лучевой и локтевой жилках, крылья остаются прямыми. У некоторых дневных видов (например, Buprestidae, Scarabaeidae), полет не включает большого подъема надкрылий, поскольку заднегрудные крылья выходят под боковые края надкрылий.[7] Высота полета жуков различна. Одно исследование по изучению высоты полета божьих коровок Coccinella septempunctata и Harmonia axyridis Использование радара показало, что, хотя большинство из них летали над одним местом на высоте 150–195 м над уровнем земли, некоторые из них достигли высоты более 1100 м.[90]
У многих бродячих жуков значительно уменьшились надкрылья, и, хотя они способны летать, они чаще всего передвигаются по земле: их мягкие тела и сильные мышцы живота делают их гибкими, легко способными извиваться в небольшие трещинки.[91]
Водные жуки используют несколько способов удержания воздуха под поверхностью воды. Жуки-ныряльщики (Dytiscidae) во время ныряния удерживают воздух между брюшком и надкрыльями. У гидрофилидов есть волосы на их внутренней поверхности, которые удерживают слой воздуха напротив их тела. Ползание взрослых водяные жуки использовать и надкрылья, и заднюю часть тазики (базальный сегмент задних ног) в задержке воздуха, а жуки-вихри просто носите с собой воздушный пузырь при каждом погружении.[92]
Коммуникация
Жуки могут общаться разными способами, в том числе с помощью феромоны. В горный сосновый жук испускает феромон, чтобы привлечь к дереву других жуков. Массы жуков способны преодолеть химическую защиту дерева. После того, как защита дерева исчерпана, жуки испускают антиагрегационный феромон. Этот вид может стричься общаться,[93] но другие могут использовать звук для защиты при нападении.[94]
Родительская забота

Родительская забота находится в нескольких семьях[95] жука, возможно, для защиты от неблагоприятных условий и хищников.[7] Бродячий жук Бледиус спектабилис живет в солончаки, поэтому яйца и личинки подвергаются опасности из-за роста прилив. Материнский жук патрулирует яйца и личинки, роя норы, чтобы они не затоплялись и удушающийи защищает их от хищного жужелица Dicheirotrichus gustavi и из паразитоидный Оса Barycnemis blediator, который убивает около 15% личинок.[96]
Жуки-закопатели являются внимательными родителями и участвуют в совместном уходе и кормлении своего потомства. Оба родителя работают, чтобы закопать тушу маленького животного, которая служит пищей для их детенышей, и построить вокруг нее выводочную камеру. Родители подготавливают тушку и защищают ее от конкурентов и от преждевременного разложения. После вылупления яиц родители защищают личинок от грибка и бактерий и помогают личинкам питаться, срыгивая для них пищу.[97]
Немного навозные жуки обеспечивать родительскую заботу, собирая помет травоядных и откладывая яйца в пределах этого корма, что является примером массовое обеспечение. Некоторые виды не уходят после этой стадии, а остаются, чтобы защитить свое потомство.[98]
Большинство видов жуков не проявляют родительского поведения после того, как откладывают яйца.[99]
Субсоциальность, когда самки охраняют свое потомство, хорошо документирована в двух семействах Chrysomelidae, Cassidinae и Chrysomelinae.[100][101][102][103][104]
Эусоциальность
Эусоциальность включает в себя совместный уход за потомством (включая уход за потомством от других особей), перекрытие поколений внутри колонии взрослых и разделение труда на репродуктивные и не репродуктивные группы.[105] Мало организмов снаружи Перепончатокрылые проявлять такое поведение; единственный жук, который так поступает, это долгоносик Austroplatypus incompertus.[106] Этот Австралийский вид обитает в горизонтальных сетях туннелей, в сердцевина из Эвкалипт деревья. Это один из более чем 300 видов сверлильных станков по дереву. Жуки-амброзии которые распространяют споры грибов амброзии.[107] Грибы растут в туннелях жуков, обеспечивая пищу жукам и их личинкам; потомство женского пола остается в туннелях и поддерживает рост грибов, вероятно, никогда не размножаясь.[107][106] Совместная забота о выводках встречается также у бессушек (Passalidae), где личинки питаются полупереваренными фекалиями взрослых особей.[108]
Кормление
_on_Ipomoea_carnea_W_IMG_0593.jpg)
Жуки могут использовать самые разные источники пищи, доступные во многих местах их обитания. Некоторые всеядные, поедая как растения, так и животных. Другие жуки очень специализированы в своем питании. Многие виды листоедов, усачей и долгоносиков очень зависят от хозяина и питаются только одним видом растений. Жужелицы и бродячие жуки (Staphylinidae), среди прочего, в первую очередь плотоядны и ловят и потребляют множество других членистоногие и мелкая добыча, такая как дождевые черви и улитки. Некоторые более крупные жуки или их личинки могут потреблять позвоночных, особенно земноводных и рыб.[109] Хотя большинство хищных жуков являются универсальными, некоторые виды имеют более специфические требования или предпочтения к добыче.[110] У некоторых видов пищеварительная способность зависит от симбиотический отношения с грибы - у некоторых жуков в кишечнике живут дрожжи, в том числе некоторые дрожжи, ранее не обнаруженные где-либо еще.[111]
Разлагающееся органическое вещество - основная пища для многих видов. Это может варьироваться от навоз, который потребляется копрофаг виды (например, определенные жуки-скарабеи в Scarabaeidae), мертвым животным, которых поедают некрофаг виды (такие как жуки-падальщики, Silphidae). Некоторые жуки, обнаруженные в навозе и падалье, на самом деле являются хищниками. В их число входят члены Histeridae и Silphidae, охотясь на личинок копрофаг и некрофаг насекомые.[112] Многие жуки питаются корой, некоторые питаются древесиной, а другие питаются грибами, растущими на древесине или опавшей листве. У некоторых жуков есть особые микангия, конструкции для переноса спор грибов.[113]
Экология

Адаптации против хищников
Жуки, как взрослые особи, так и личинки, являются добычей многих животных. хищники включая млекопитающие из летучие мыши к грызуны, птицы, ящерицы, амфибии, Рыбы, стрекозы, грабители, повторяющиеся ошибки, муравьи, другие жуки и пауки.[114][115] Жуки используют различные антихищные приспособления защищаться. К ним относятся камуфляж и мимикрия против хищников, которые охотятся на вид, токсичность и защитное поведение.
Камуфляж
Камуфляж обычен и широко распространен среди семейств жуков, особенно тех, которые питаются древесиной или растительностью, например листоеды (Chrysomelidae, часто зеленые) и долгоносики. У некоторых видов скульптура или разноцветные чешуйки или волоски вызывают появление жуков, таких как авокадо долгоносик Heilipus apiatus напоминать птичий помет или другие несъедобные предметы.[114] Многие жуки, обитающие в песчаной среде, сливаются с окраской этого субстрата.[116]
Мимикрия и апосематизм

Немного усачьи жуки (Cerambycidae) эффективны Бейтсовская имитация из осы. Жуки могут сочетать окраску с поведенческой мимикой, действуя как осы, на которых они уже очень похожи. Многие другие жуки, в том числе божьи коровки, волдыри, и лицидные жуки выделяют неприятные или ядовитые вещества, чтобы сделать их неприятными или ядовитыми, и часто апосематический, где яркие или контрастные окраска предупредить хищников; многие жуки и другие насекомые имитируют эти химически защищенные виды.[117]
.jpg)

Химическая защита важна для некоторых видов и обычно рекламируется яркими апосематическими цветами. Немного Tenebrionidae используют свою позу для выпуска ядовитых химикатов, чтобы предупредить хищников. Химическая защита может служить не только для защиты от позвоночных, но и для других целей, например, для защиты от широкого спектра микробов. Некоторые виды выделяют химические вещества из растений, которыми питаются, и включают их в свою защиту.[116]
У других видов есть специальные железы для производства отпугивающих химикатов. Защитные железы жужелиц-жужелиц производят различные углеводороды, альдегиды, фенолы, хиноны, сложные эфиры, и кислоты выпущен из отверстия в конце живота. Африканские жужелицы (например, Anthia) используют те же химические вещества, что и муравьи: муравьиная кислота.[117] Жуки-бомбардиры имеют хорошо развитые пигидиальные железы, которые опорожняются с боков межсегментных перепонок между седьмым и восьмым брюшными сегментами. Сальник состоит из двух вмещающих камер, одна для гидрохиноны и пероксид водорода, другой - перекись водорода и каталаза ферменты. Эти химические вещества смешиваются и приводят к взрывному выбросу, достигающему температуры около 100 ° C (212 ° F), с распадом гидрохинона на водород, кислород и хинон. Кислород запускает ядовитые химические брызги как струю, которая может быть точно нацелена на хищников.[7]
Другая защита
Крупные наземные жуки, такие как Carabidae, то Жук-носорог а усачьи защищаются сильными челюсти, или сильно склеротизированные (бронированные) шипы или рога, чтобы отпугивать или отбиваться от хищников.[116] Многие виды долгоносиков, которые питаются на открытом воздухе листьями растений, реагируют на нападение с помощью рефлекса опускания. Некоторые сочетают это с танатоз, в котором они закрывают свои придатки и «притворяются мертвыми».[118] Жуки-щелкуны (Elateridae) могут внезапно катапультироваться, спасаясь от опасности, высвобождая энергию, запасенную механизмом щелчка, который состоит из толстого позвоночника на переднегруди и соответствующей бороздки в мезостерне.[114] Некоторые виды пугают атакующего, издавая звуки с помощью процесса, известного как скрежетание.[94]
Паразитизм
Несколько видов жуков эктопаразитарный на млекопитающих. Один из таких видов, Platypsyllus castoris, паразитирует на бобрах (Кастор виды). Этот жук живет как паразит как личинка, так и взрослый человек, питаясь эпидермальной тканью и, возможно, кожными выделениями и экссудатами из ран. Они поразительно сплюснуты дорсовентрально, несомненно, как приспособление для скольжения между бобровыми волосками. Они бескрылые и безглазые, как и многие другие эктопаразиты.[119] Другие являются клептопаразитами других беспозвоночных, таких как маленький улей (Aethina tumida) который заражает пчела гнезда[120] в то время как многие виды паразитируют инквилинс или же комменсал в муравейниках.[121] Несколько групп жуков являются первичными паразитоиды других насекомых, питающихся и в конечном итоге убивающих своих хозяев.[122]
Опыление

Цветки, опыляемые жуками, обычно большие, зеленоватого или кремового цвета и сильно пахнут. Ароматы могут быть пряными, фруктовыми или похожими на запах разлагающихся органических веществ. Жуки, скорее всего, были первыми насекомыми, которые опыляли цветы.[нужна цитата] Большинство опыляемых жуками цветов имеют уплощенную или чашевидную форму с легкодоступной пыльцой, хотя они могут включать ловушки держать жука подольше. Завязи растений обычно хорошо защищены от кусания ротовой полости их опылителей. Семейства жуков, которые обычно опыляют цветы, являются Buprestidae, Cantharidae, Cerambycidae, Cleridae, Dermestidae, Lycidae, Мелирид, Морделлиды, Nitidulidae и Scarabaeidae.[123] Жуки могут быть особенно важны в некоторых частях мира, таких как полузасушливые районы юга Африки и Южная Калифорния[124] и горные луга Квазулу-Натал в Южной Африке.[125]
Мутуализм

2: Личинка питается грибком, который переваривает древесину, удаляя токсины, к взаимной выгоде.
3: Личинка окукливается.
Мутуализм хорошо известен у некоторых жуков, таких как жук-амброзия, который вместе с грибами переваривает древесину мертвых деревьев. Жуки роют туннели в мертвых деревьях, где выращивают грибные сады, единственный источник питания. После приземления на подходящее дерево жук-амброзия выкапывает туннель, в котором он выпускает споры своего грибка. симбионт. Гриб проникает в ткань ксилемы растения, переваривает ее и концентрирует питательные вещества на поверхности галереи жуков и вблизи нее, так что долгоносики и гриб получают пользу. Жуки не могут есть древесину из-за токсинов и используют свои отношения с грибами, чтобы помочь преодолеть защиту своего дерева-хозяина, чтобы обеспечить питание для своих личинок.[126] Химически опосредован полиненасыщенным пероксидом, продуцируемым бактериями,[127] эта мутуалистическая связь между жуком и грибом совместно развитый.[126][128]

Устойчивость к экстремальным условиям окружающей среды

Около 90% видов жуков вступают в период взросления. диапауза, спокойная фаза с пониженным метаболизмом, чтобы избежать неблагоприятных условий окружающей среды. Диапауза взрослых особей - наиболее частая форма диапаузы жесткокрылых. Чтобы выдержать период без еды (часто длящийся много месяцев), взрослые готовятся, накапливая запасы липидов, гликогена, белков и других веществ, необходимых для устойчивости к будущим опасным изменениям условий окружающей среды. Эта диапауза вызвана сигналами, предвещающими наступление неблагоприятного сезона; обычно реплика фотопериодический. Короткий (уменьшающийся) световой день служит сигналом о приближении зимы и вызывает зимнюю диапаузу (гибернацию).[129] Исследование спячки арктического жука Pterostichus brevicornis показали, что уровень жира в организме взрослых был самым высоким осенью с пищеварительный тракт заполнены едой, но к концу января опустеют. Эта потеря жира в организме была постепенным процессом, происходящим в сочетании с обезвоживанием.[130]
Все насекомые пойкилотермный,[131] поэтому способность некоторых жуков жить в экстремальных условиях зависит от их устойчивости к необычно высоким или низким температурам. В короед Pityogenes chalcographus может выжить −39° C во время зимовки под корой дерева;[132] аляскинский жук Cucujus clavipes puniceus способен выдержать −58° C; его личинки могут выжить −100° C.[133] При таких низких температурах образование кристаллов льда во внутренних жидкостях является самой большой угрозой выживанию жуков, но этому препятствует выработка белков-антифризов, которые не позволяют молекулам воды группироваться вместе. Низкие температуры, испытанные Cucujus clavipes могут выжить благодаря их преднамеренному обезвоживанию в сочетании с антифризами. Это в несколько раз концентрирует антифриз.[134] В гемолимфа жука мучного червя Тенебрио Молитор содержит несколько антифризы протеины.[135] Аляскинский жук Upis ceramboides выдерживает −60 ° C: его криопротекторы находятся ксиломаннан, молекула, состоящая из сахар привязан к жирная кислота,[136] и сахар-спирт, треитол.[137]
И наоборот, пустынные жуки приспособлены к высоким температурам. Например, Тенебрионид жук Onymacris rugatipennis может выдержать 50° C.[138] Тигровые жуки в жарких песчаных районах часто бывают белесыми (например, Habroscelimorpha dorsalis), чтобы отразить больше тепла, чем более темный цвет. Эти жуки также демонстрируют поведенческую адаптацию, чтобы переносить жару: они могут стоять прямо на лапках, чтобы удерживать свое тело от горячей земли, искать тень и поворачиваться лицом к солнцу, так что только передние части их головы находятся прямо незащищенный.[139]
Жук тумана Пустыня Намиб, Stenocara gracilipes, способен собирать воду из тумана, так как его надкрылья имеют текстурированную поверхность, сочетающую гидрофильный (водолюбивые) шишки и восковые, гидрофобный кормушки. Жук встречает утренний ветерок, приподнимая брюшко; капли конденсируются на надкрыльях и бегут по гребням к ротовому аппарату. Подобные приспособления обнаружены у некоторых других жуков пустыни Намиб, таких как Onymacris unguicularis.[140]
Некоторые наземные жуки, эксплуатирующие береговую линию и места обитания в поймах, обладают физиологической адаптацией, чтобы пережить наводнения. В случае наводнения взрослые жуки могут быть достаточно мобильными, чтобы уйти от наводнения, но личинки и куколки часто не могут. Взрослые Cicindela togata не могут пережить погружение в воду, но личинки способны выжить в течение длительного периода, до 6 дней, аноксия во время паводков. Толерантность к аноксии личинок могла поддерживаться переключением на анаэробные метаболические пути или снижением скорости метаболизма.[141] Переносимость аноксии у взрослых жужелиц Пелофилия borealis был протестирован в лабораторных условиях, и было обнаружено, что они могут выдерживать непрерывный период до 127 дней в атмосфере 99,9% азота при 0 ° C.[142]
Миграция
Многие виды жуков совершают ежегодные массовые перемещения, которые называются миграциями. К ним относятся пыльцевый жук Meligethes aeneus[143] и многие виды кокцинеллиды.[144] Эти массовые движения также могут быть оппортунистическими в поисках пищи, а не сезонными. Исследование 2008 г. необычно крупной вспышки горного соснового жука (Dendroctonus ponderosae) в британская Колумбия обнаружили, что жуки способны пролетать 30–110 км в день при плотности до 18 600 жуков на гектар.[145]
Отношение к людям
В древних культурах
| ||
| ḫpr в иероглифы |
|---|

Несколько видов навозных жуков, особенно священный скарабей, Скарабей-жрец, почитались в Древний Египет.[146][147] Иероглифический образ жука мог иметь экзистенциальное, вымышленное или онтологическое значение.[148] Изображения скарабея в кости, слоновая кость, камень, Египетский фаянс, и драгоценные металлы известны от Шестой династии до периода римского правления. Скарабей имел первостепенное значение в погребальном культе Древнего Египта.[149] Скарабей был связан с Хепри, бог восхода солнце, от предполагаемого сходства катания навозного комка жуком с катанием солнца богом.[146] Некоторые соседи Древнего Египта переняли мотив скарабея для уплотнения разных типов. Самыми известными из них являются иудейские Уплотнения ЛМЛК, где восемь из 21 рисунка содержали жуков-скарабеев, которые использовались исключительно для штамповки оттисков на сосудах для хранения во время правления Езекия.[150] Жуки упоминаются как символ солнца, как в Древнем Египте, в Плутарх1 век Мораль.[151] В Греческие волшебные папирусы от 2-го века до нашей эры до 5-го века нашей эры описывают скарабеев как ингредиент заклинания.[152]
Плиний Старший обсуждает жуков в своем Естественная история,[153] описывая жук-олень: "Некоторых насекомых для сохранения своих крыльев покрывают эрстом (надкрылья) - например, жук, у которого особенно тонкое и хрупкое крыло. Природа отказала этим насекомым в укусе; но у одного большого вида мы находим рога замечательной длины, с двумя зубцами на концах и образующими клешни, которые животное закрывает, когда собирается укусить ».[154][155] Жук-олень описан в греческом мифе. Никандер и напомнил Антонин Либералис в котором Церамбус[b] превращается в жука: «Его можно увидеть на стволах, и у него есть крючковатые зубы, он постоянно двигает челюстями вместе. Он черный, длинный и с твердыми крыльями, как у большого навозного жука».[156] История завершается комментарием о том, что мальчики использовали жуков в качестве игрушек, а голову снимали и носили как подвеску.[155][157]
Как вредители

Около 75% видов жуков являются фитофагами как на личиночной, так и на взрослой стадии. Многие питаются экономически важными растениями и хранимыми растительными продуктами, включая деревья, злаки, табак и сухофрукты.[6] Некоторые, например долгоносик, который питается ватными палочками и цветами, может нанести чрезвычайно серьезный ущерб сельскому хозяйству. Долгоносик пересек Рио-Гранде возле Brownsville, Техас, чтобы войти в Соединенные Штаты из Мексика около 1892 г.,[158] и достиг юго-востока Алабама к 1915 году. К середине 1920-х годов он проник во все хлопковые регионы США, путешествуя от 40 до 160 миль (60–260 км) в год. Он остается самым разрушительным вредителем хлопка в Северной Америке. Государственный университет Миссисипи По оценкам, с тех пор, как долгоносик попал в Соединенные Штаты, он обходился производителям хлопка примерно в 13 миллиардов долларов, а в последнее время - примерно в 300 миллионов долларов в год.[158]
Короед, ильм листоед и азиатский усачок (Anoplophora glabripennis)[159] относятся к числу видов, нападающих вяз деревья. Короеды (Scolytidae) нести Болезнь голландского вяза когда они переходят с зараженных мест размножения на здоровые деревья. Болезнь поразила ильмы по всей Европе и Северной Америке.[160]

У некоторых видов жуков выработался иммунитет к инсектицидам. Например, Колорадский жук, Leptinotarsa decemlineata, является истребителем растений картофеля. Среди его хозяев - другие члены Пасленовые, Такие как паслен, помидор, баклажан и стручковый перец, а также картофель. У разных популяций выработалась устойчивость ко всем основным классам инсектицидов.[161] Колорадский жук оценивался как орудие энтомологическая война в течение Вторая Мировая ВойнаИдея состоит в том, чтобы использовать жука и его личинок, чтобы повредить урожай вражеских народов.[162] Германия протестировала свою программу создания оружия против картофельного жука к югу от Франкфурт, выпустив 54 000 жуков.[163]
В жук часы смерти, Xestobium rufovillosum (Ptinidae), является серьезным вредителем старых деревянных построек в Европе. Он атакует лиственных пород Такие как дуб и каштан, всегда там, где произошел или имеет место грибковый распад. Считается, что фактическая интродукция вредителя в здания происходит во время строительства.[164]
К другим вредителям относятся кокосовый гистолог, Бронтиспа лонгиссима, который питается молодыми листья, саженцы и зрелый кокос деревья, нанесшие серьезный экономический ущерб Филиппины.[165] В горный сосновый жук является разрушительным вредителем зрелых или ослабленных Ложная сосна, иногда поражая большие территории Канады.[166]
Как полезные ресурсы

Жуки могут быть полезны для экономики человека, контролируя популяции вредителей. Личинки и имаго некоторых видов божьи коровки (Coccinellidae) питаться тля это вредители. Другие жуки-беди поедают щитовки, белокрылка и мучнистые червецы.[167] Если нормальных источников пищи мало, они могут питаться небольшими гусеницы, молодой жуки растений, или же нектар и нектар.[168] Жужелицы (Carabidae) обычны хищники многих насекомых-вредителей, включая яйца мух, гусениц и проволочников.[169] Жужелицы могут помочь контролировать сорняки поедая их семена в почве, уменьшая потребность в гербициды для защиты посевов.[170] Эффективность некоторых видов в сокращении определенных популяций растений привела к преднамеренной интродукции жуков для борьбы с сорняками. Например, род Зигограмма родом из Северной Америки, но использовался для контроля Парфений истерофор в Индии и Амброзия полыннолистная в России.[171][172]
Навозные жуки (Scarabidae) успешно использовались для сокращения популяций вредных мух, таких как Musca vetustissima и Haematobia exigua которые являются серьезными вредителями крупного рогатого скота в Австралия.[173] Жуки делают помет недоступным для размножающихся вредителей, быстро перекатывая и закапывая его в почву, с дополнительным эффектом улучшения плодородия почвы, вспашки и круговорота питательных веществ.[174] В Австралийский проект по уничтожению навозных жуков (1965–1985), ввел в Австралию виды навозных жуков из Южной Африки и Европы для сокращения популяций Musca vetustissima, после успешных испытаний этой техники в Гавайи.[173] В Американский институт биологических наук сообщает, что навозные жуки спасают животноводство Соединенных Штатов примерно на 380 миллионов долларов США ежегодно за счет захоронения фекалий домашнего скота над землей.[175]
В Dermestidae часто используются в таксидермия и при подготовке научных образцов, чтобы очистить мягкие ткани от костей.[176] Личинки питаются и удаляют хрящи вместе с другими мягкими тканями.[177][178]
Как еда и лекарства

Жуки - наиболее часто употребляемые в пищу насекомые, их в пищу употребляют около 344 видов, обычно на стадии личинок.[179] В хрущак мучной (личинка чернотелка) и Жук-носорог относятся к числу обычно употребляемых в пищу видов.[180] Широкий спектр видов также используется в Народная медицина для лечения людей, страдающих различными расстройствами и заболеваниями, хотя это делается без клинических исследований, подтверждающих эффективность такого лечения.[181]
Как индикаторы биоразнообразия
Из-за специфики среды обитания многие виды жуков были предложены в качестве подходящих индикаторов, их присутствие, численность или отсутствие, позволяющее измерить качество среды обитания. Хищные жуки, такие как тигровые жуки (Cicindelidae) нашли научное применение в качестве таксона-индикатора для измерения региональных структур биоразнообразия. Они подходят для этого, поскольку их систематика стабильна; история их жизни хорошо описана; они большие, и их легко наблюдать при посещении сайта; они встречаются по всему миру во многих средах обитания, причем виды специализируются на определенных средах обитания; и их встречаемость по видам точно указывает на другие виды, как позвоночные, так и беспозвоночные.[182] Судя по местам обитания, многие другие группы, такие как бродячие жуки в измененных человеком средах обитания, навозные жуки в саваннах[183] и сапроксильных жуков в лесах[184] были предложены в качестве потенциальных индикаторных видов.[185]
В искусстве и украшениях


У многих жуков красивые и прочные надкрылья, которые использовались в качестве материала в искусстве. жук лучший пример.[186] Иногда они включаются в ритуальные предметы из-за их религиозного значения. Из целых жуков в чистом виде или в прозрачной пластмассовой оболочке превращаются в различные предметы, от дешевых сувениров, таких как брелки, до дорогих ювелирных украшений. В некоторых частях Мексики жуки этого рода Zopherus превращены в живые броши прикрепляя бижутерию и золотые цепочки, что стало возможным благодаря невероятно твердым надкрыльям и малоподвижному образу жизни этого рода.[187]
В развлечениях
Боевые жуки используются для развлечений и азартных игр. Этот вид спорта использует территориальное поведение и спаривание определенных видов крупных жуков. в Чиангмай район северного Таиланда, мужской Ксилотрупы Жуков-носорогов ловят в дикой природе и дрессируют для борьбы. Самок держат внутри бревна, чтобы стимулировать борющихся самцов своими феромонами.[188] Эти бои могут быть соревновательными и включать в себя азартные игры как деньгами, так и имуществом.[189] В Южная Корея то Dytiscidae разновидность Cybister tripunctatus используется в игре, подобной рулетке.[190]
Жуков иногда используют в качестве инструментов: Онабасулу из Папуа - Новая Гвинея исторически использовался долгоносик Rhynchophorus ferrugineus как музыкальный инструмент, позволяя человеческому рту служить переменной резонансная камера для колебаний крыльев взрослого живого жука.[189]
Как домашние животные
Некоторые виды жуков содержатся как домашние питомцы, например ныряльщики (Dytiscidae) можно хранить в резервуаре для бытовой пресной воды.[191]

В Япония практика содержания рогатых жуков-носорогов (Dynastinae) и жуков-оленей (Lucanidae) особенно популярен среди мальчиков.[192] Популярность в Японии такова, что торговые автоматы В 1999 году были выведены раздаточные животы, в каждом из которых содержится до 100 особей-оленей.[193][194]
Как вещи собирать
Коллекционирование жуков стало чрезвычайно популярным в Викторианская эпоха.[195] Натуралист Альфред Рассел Уоллес собрал (по его собственным подсчетам) в общей сложности 83 200 жуков за восемь лет, описанных в его книге 1869 года. Малайский архипелаг, в том числе 2000 новых для науки видов.[196]
Как вдохновение для технологий
Некоторые приспособления жесткокрылых вызвали интерес у биомиметика с возможными коммерческими приложениями. В Жук-бомбардирМощный репеллентный спрей вдохновил на разработку технологии распыления мелкодисперсного тумана, который, как утверждается, имеет низкое углеродное воздействие по сравнению с аэрозольными распылителями.[197] Уборка влаги пустынным жуком Намиб (Stenocara gracilipes) вдохновил на создание самонаполняющейся бутылки для воды, в которой гидрофильный и гидрофобный материалы для людей, живущих в засушливых регионах без регулярных дождей.[198]
Живые жуки использовались как киборги. А Агентство перспективных оборонных исследовательских проектов финансируемый проект имплантировал электроды в Mecynorhina torquata жуков, позволяя дистанционно управлять ими с помощью радиоприемника, удерживаемого на его спине, в качестве доказательства концепции для работы по наблюдению.[199] Аналогичная технология была применена, чтобы позволить человеку-оператору управлять свободным полетом и походкой Mecynorhina torquata а также ступенчатый поворот и обратная ходьба Zophobas morio.[200][201][202]
Исследование, опубликованное в 2020 году, было направлено на создание робота-рюкзака для жуков. Миниатюрные фотоаппараты весом 248 мг прикреплялись к живым жукам Тенебрионид роды Асболус и Элеоды. Камеры снимали под углом 60 ° до 6 часов.[203][204]
В консервации
Поскольку жуки составляют значительную часть мирового биоразнообразия, их сохранение имеет важное значение, и, в равной степени, потеря среды обитания и биоразнообразия, несомненно, повлияет на жуков. Многие виды жуков имеют очень специфические среды обитания и длительный жизненный цикл, что делает их уязвимыми. Некоторые виды находятся под серьезной угрозой, в то время как другие уже находятся под угрозой исчезновения.[205] Островные виды, как правило, более восприимчивы, как в случае Helictopleurus undatus Мадагаскара, который, как считается, вымер в конце 20 века.[206] Защитники природы пытались пробудить симпатию к жукам с помощью флагманских видов, таких как жук-олень, Lucanus cervus,[207] и тигровые жуки (Cicindelidae). В Японии светлячок Гэндзи, Luciola cruciata, чрезвычайно популярен, а в Южной Африке слоновый навозник Аддо предлагает многообещающие возможности для расширения экотуризм за пределами Большая пятерка туристических видов млекопитающих. Популярная неприязнь к жукам-вредителям тоже может превратиться в общественный интерес к насекомым, как и необычная экологическая адаптация таких видов, как фея-креветка-охотничий жук. Cicinis bruchi.[208]
Примечания
Рекомендации
- ^ а б Bouchard, P .; Bousquet, Y .; Дэвис, А .; Алонсо-Саразага, М .; Lawrence, J .; Lyal, C .; Ньютон, А .; Reid, C .; Schmitt, M .; Слипинский, А .; Смит, А. (2011). "Названия группы семейства жесткокрылых (Insecta)". ZooKeys (88): 1–972. Дои:10.3897 / zookeys.88.807. ЧВК 3088472. PMID 21594053.
- ^ Харпер, Дуглас. «Жесткокрылые». Интернет-словарь этимологии.
- ^ Харпер, Дуглас. "Жук". Интернет-словарь этимологии.
- ^ "Жук". Онлайн-словарь Merriam-Webster. Получено 20 февраля, 2016.
- ^ Харпер, Дуглас. "Чефер". Интернет-словарь этимологии.
- ^ а б c d е ж грамм час я Гиллиотт, Седрик (август 1995). Энтомология (2-е изд.). Springer-Verlag. п. 96. ISBN 978-0-306-44967-3.
- ^ а б c d е ж грамм час я j k л м п о п q МакХью (2009)
- ^ Розенцвейг, М. Л. (1995). Разнообразие видов в пространстве и времени. Кембридж: Издательство Кембриджского университета. п. 2. ISBN 978-0-521-49952-1.
- ^ а б Хант, Т .; Bergsten, J .; Левканикова, З .; Papadopoulou, A .; John, O.S .; Wild, R .; Hammond, P.M .; Аренс, Д .; Балке, М .; Катерино, М. С .; Gómez-Zurita, J .; Рибера, I; Barraclough, T. G .; Бочакова, М .; Bocak, L; Фоглер, А. П. (2007). «Всеобъемлющая филогения жуков раскрывает эволюционное происхождение сверхизлучения». Наука. 318 (5858): 1913–1916. Bibcode:2007Научный ... 318.1913H. Дои:10.1126 / science.1146954. PMID 18096805. S2CID 19392955.
- ^ Хаммонд, П. (1992). Инвентаризация видов. Глобальное биоразнообразие, состояние живых ресурсов Земли: отчет (1-е изд.). Лондон: Чепмен и Холл. стр.17–39. ISBN 978-0-412-47240-4.
- ^ а б Аист, Найджел Э .; Макбрум, Джеймс; Гели, Клэр; Гамильтон, Эндрю Дж. (2015). «Новые подходы сужают глобальные оценки видов жуков, насекомых и наземных членистоногих». PNAS. 116 (24): 7519–7523. Bibcode:2015ПНАС..112.7519С. Дои:10.1073 / pnas.1502408112. ЧВК 4475949. PMID 26034274.
- ^ Gullan, P.J .; Крэнстон, П.С. (2014). Насекомые: очерк энтомологии (5-е изд.). Джон Вили и сыновья. п.6. ISBN 978-1-4443-3036-6.
- ^ Gullan, P.J .; Крэнстон, П. С. (2014). Насекомые: очерк энтомологии (5-е изд.). Джон Вили и сыновья. п.517. ISBN 978-1-4443-3036-6.
- ^ Кирмс С., Адис Дж., Моравец В. 2003. События цветения и разнообразие жуков в Венесуэле. В: Бассет Ю., Новотный В., Миллер С.Е., Китчинг Р.Л., редакторы. Членистоногие тропических лесов: пространственно-временная динамика и использование ресурсов в пологе. Кембридж: Издательство Кембриджского университета; п. 256–265.
- ^ Арндт, Эрик; Кирмс, Сьюзен; Эрвин, Терри Л. (2001). «Древесные жуки неотропических лесов: Агра. Фабрициус, Описания личинок с примечаниями по естествознанию и поведению (Coleoptera, Carabidae, Lebiini, Agrina) ». Бюллетень колеоптерологов. 55 (3): 297–310. Дои:10.1649 / 0010-065x (2001) 055 [0297: abonfa] 2.0.co; 2.
- ^ Кирмс, Сьюзен; Чабу, Кэролайн С. (2018). «В сообществе листоедов (Coleoptera: Chrysomelidae), населяющих полог тропических равнинных тропических лесов на юге Венесуэлы, преобладают полифагия и флористика». Журнал естественной истории. 52 (41–42): 2677–2721. Дои:10.1080/00222933.2018.1548666. S2CID 91732501.
- ^ Кирмс, Сьюзен; Рэтклифф, Бретт С. (2019). «Состав и образцы использования хозяев сообщества жука-скарабея (Coleoptera: Scarabaeidae), населяющего полог тропических лесов на низменности в южной части Венесуэлы». Бюллетень колеоптерологов. 73: 149. Дои:10.1649 / 0010-065X-73.1.149. S2CID 108786139.
- ^ «Самое тяжелое насекомое». Книга Рекордов Гиннесса. Получено 1 февраля, 2017.
- ^ Уильямс, Дэвид М. (2001). «Глава 30 - Самое большое насекомое». Книга записей о насекомых. Университет Флориды. Архивировано из оригинал 18 июля 2011 г.
- ^ Полилов, Алексей (2015). «Насколько маленькое самое маленькое? Новый рекорд и повторные измерения Scydosella musawasensis Hall, 1999 (Coleoptera, Ptiliidae), самого маленького из известных свободноживущих насекомых». ZooKeys (526): 61–64. Дои:10.3897 / zookeys.526.6531. ЧВК 4607844. PMID 26487824.
- ^ а б c d е ж грамм час Бениш, Кристоф (2010). «Филогения жуков». Фауна жуков Германии. Кербтье. Получено 16 марта, 2011.
- ^ Гримальди, Дэвид; Энгель, Майкл С. (2005). Эволюция насекомых. Издательство Кембриджского университета. п. 397. ISBN 978-1-107-26877-7.
Триасовые Curculionoidea имеют тот же статус, что и триасовые Chrysoleloidea: связь, которая крайне маловероятна и в настоящее время невозможно проверить.
- ^ Бету, Оливер (2009). «Самый ранний идентифицированный жук». Журнал палеонтологии. 83 (6): 931–937. Дои:10.1666/08-158.1. S2CID 85796546.
- ^ Hörnschemeyer, T .; Стапф, Х. "Die Insektentaphozönose von Niedermoschel (Асселиан, ун. Пермь; Германия)". Schriften der Alfred-Wegener-Stiftung (на немецком языке) (99/8): 98.
- ^ Кукалова, J. (1969). «О систематическом положении предполагаемых пермских жуков, Tshecardocoleidae, с описанием новой коллекции из Моравии». Sborník Geologických Věd, Paleontologie. 11: 139–161.
- ^ Beckemeyer, R.J .; Энгель, М. С. (2008). "Второй образец Пермоколей (Coleoptera) из нижнепермской формации Веллингтон округа Нобл, Оклахома " (PDF). Журнал Канзасского энтомологического общества. 81 (1): 4–7. Дои:10.2317 / JKES-708.01.1. S2CID 86835593. Архивировано из оригинал (PDF) 18 июля 2011 г.
- ^ Щербаков, Д. Э. (2008). «О фауне насекомых перми и триаса в связи с биогеографией и пермско-триасовым кризисом». Палеонтологический журнал. 42 (1): 15–31. Дои:10.1007 / s11492-008-1003-1. S2CID 128919393.
- ^ Пономаренко, А. Г. (2004). «Жуки (Insecta, Coleoptera) поздней перми и раннего триаса» (PDF). Палеонтологический журнал. 38 (Приложение 2): S185 – S196. Архивировано из оригинал (PDF) 11 ноября 2013 г.
- ^ а б c МакХью (2009), п. 186
- ^ Labandeira, C.C .; Сепкоски, Дж. Дж. (1993). «Разнообразие насекомых в летописи окаменелостей» (PDF). Наука. 261 (5119): 310–315. Bibcode:1993Наука ... 261..310Л. CiteSeerX 10.1.1.496.1576. Дои:10.1126 / science.11536548. PMID 11536548. Архивировано из оригинал (PDF) 31 марта 2012 г.
- ^ Грачев, Вадим Г .; Жерихин Владимир Васильевич (15 октября 2003 г.). «Разнообразие насекомых в летописи окаменелостей» (PDF). Acta Zoologica Cracoviensia. 261 (5119): 129–138. Bibcode:1993Научный ... 261..310Л. CiteSeerX 10.1.1.496.1576. Дои:10.1126 / science.11536548. PMID 11536548.
- ^ Chang, H .; Zhang, F .; Рен, Д. (2008). «Новый род и два новых вида ископаемых элатерид из формации Исянь в Западном Ляонине, Китай (Coleoptera: Elateridae)» (PDF). Zootaxa. 1785 (1): 54–62. Дои:10.11646 / zootaxa.1785.1.3. Архивировано из оригинал (PDF) 4 июля 2011 г.
- ^ Орехово-Зуево, А.В.А. (1993). «Юрские и нижнемеловые Buprestidae (Coleoptera) из Евразии» (PDF). Палеонтологический журнал (1A): 9–34.
- ^ Пономаренко, Александр Г. (1985). «Ископаемые насекомые из титонского 'Solnhofener Plattenkalke' в Музее естественной истории, Вена» (PDF). Annalen des Naturhistorischen Museums в Вене. 87 (1): 135–144.
- ^ Ян, Э. В. (2009). «Новый род жукообразных жуков (Coleoptera, Polyphaga) из средней-поздней юры Каратау» (PDF). Палеонтологический журнал. 43 (1): 78–82. Дои:10.1134 / S0031030109010080. S2CID 84621777. Архивировано из оригинал (PDF) 18 июля 2011 г.
- ^ а б Tan, J.-J .; Ren, D .; Лю М. (2005). «Новые омматиды из поздней юры западной части Ляонина, Китай (Coleoptera: Archostemata)» (PDF). Наука о насекомых. 12 (3): 207–216. Дои:10.1111 / j.1005-295X.2005.00026.x. Архивировано из оригинал (PDF) 18 июля 2011 г.
- ^ Пономаренко, А.Г. (1997). «Новые жуки семейства Cupedidae из мезозоя Монголии. Ommatini, Mesocupedini, Priacmini» (PDF). Палеонтологический журнал. 31 (4): 389–399. Архивировано из оригинал (PDF) 25 сентября 2006 г.
- ^ Алексеев, А. В. (2009). «Новые жуки-драгоценности (Coleoptera: Buprestidae) из мелового периода России, Казахстана и Монголии» (PDF). Палеонтологический журнал. 43 (3): 277–281. Дои:10.1134 / s0031030109030058. S2CID 129618839. Архивировано из оригинал (PDF) 18 июля 2011 г.
- ^ Чин, К .; Гилл, Б. Д. (1996). «Динозавры, навозные жуки и хвойные деревья; участники пищевой сети мелового периода». ПАЛАИ. 11 (3): 280–285. Bibcode:1996 Палай..11..280C. Дои:10.2307/3515235. JSTOR 3515235.
- ^ Арилло, Антонио и Ортуньо, Висенте М. (2008). «Имели ли динозавры какое-либо отношение к навозным жукам? (Происхождение копрофагии)». Журнал естественной истории. 42 (19&20): 1405–1408. Дои:10.1080/00222930802105130. S2CID 83643794.
- ^ Дэвид Перис; Джес Руст (2020). «Меловые жуки (Insecta: Coleoptera) в янтаре: палеоэкология этой самой разнообразной группы насекомых». Зоологический журнал Линнеевского общества. 189 (4): 1085–1104. Дои:10.1093 / zoolinnean / zlz118.
- ^ McNamara, M.E .; Briggs, D. E. G .; Орр, П. Дж .; Но, H .; Цао, Х. (2011). «Оригинальные окрасы ископаемых жуков». Труды Королевского общества B: биологические науки. 279 (1731): 1114–1121. Дои:10.1098 / rspb.2011.1677. ЧВК 3267148. PMID 21957131.
- ^ Купе, Г. Р. (1979). «Позднекайнозойские ископаемые жесткокрылые: эволюция, биогеография и экология». Ежегодный обзор экологии и систематики. 10: 246–267. Дои:10.1146 / annurev.es.10.110179.001335. JSTOR 2096792.
- ^ Хатчинсон, Г. Э. (1959). «Посвящение Святой Розалии или почему так много видов животных?». Американский натуралист. 93 (870): 145–159. Дои:10.1086/282070. JSTOR 2458768.
- ^ Мэддисон, Д. Р. (1995). «Полифага». Веб-проект "Древо жизни". Получено 27 февраля, 2016.
- ^ Beutel, R.G .; Лоуренс, Дж. Ф. (2005). «4. Жесткокрылые (морфология)». In Kristensen, N.P .; Beutel, R.G. (ред.). Справочник по зоологии, группа 4: членистоногие, 2: насекомые, жесткокрылые, жуки. Том 1: Морфология и систематика (Archostemata, Adephaga, Myxophaga, Polyphaga partim). п. 23. ISBN 978-3-11-017130-3.
- ^ Beutel, R.G .; Рибера, И. (2005). «7. Адефага Шелленберг, 1806». In Kristensen, N.P .; Beutel, R.G. (ред.). Справочник по зоологии, группа 4: членистоногие, 2: насекомые, жесткокрылые, жуки. Том 1: Морфология и систематика (Archostemata, Adephaga, Myxophaga, Polyphaga partim). п. 54. ISBN 978-3-11-017130-3.
- ^ "Подотряд Archostemata - сетчатые и телефонные жуки". BugGuide. 2006 г.. Получено 26 января, 2017.
- ^ а б c Лоуренс, Джон Ф .; Слипинский, Адам (2013). Австралийские жуки. Морфология, классификация и ключи. CSIRO. С. 1–16. ISBN 978-0-643-09728-5.
- ^ Месарос, Габор (2013). «Sphaeriusidae (Coleoptera, Myxophaga): новое семейство жуков в фауне Сербии». Вестник Музея естественной истории (6): 71–74. Дои:10.5937 / bnhmb1306071m.
- ^ Beutel, Rolf G .; Лешен, Ричард (2005). Справочник по зоологии. Том 4. Часть 38. Членистоногие. Морфология и систематика (Archostemata, Adephaga, Myxophaga, Polyphaga partim). Вальтер де Грюйтер. п. 43.
- ^ Уайтинг, Майкл Ф. (2002). «Филогения отрядов голометаболических насекомых: молекулярные доказательства». Zoologica Scripta. 31 (1): 3–15. Дои:10.1046 / j.0300-3256.2001.00093.x.
- ^ Beutel, R .; Хаас, Ф. (2000). «Филогенетические взаимоотношения подотрядов Coleoptera (Insecta)». Кладистика. 16: 103–141. Дои:10.1111 / j.1096-0031.2000.tb00350.x.
- ^ а б Kukalová-Peck, J .; Лоуренс, Дж. Ф. (1993). «Эволюция заднего крыла жесткокрылых». Канадский энтомолог. 125 (2): 181–258. Дои:10.4039 / Ent125181-2.
- ^ Мэддисон, Д. Р .; Мур, В .; Baker, M.D .; Ellis, T. M .; Обер, К. А .; Cannone, J. J .; Гутелл, Р. Р. (2009). «Монофилия наземных жуков-адефаганов, на что указывают три ядерных гена (Coleoptera: Carabidae и Trachypachidae)». Zoologica Scripta. 38 (1): 43–62. Дои:10.1111 / j.1463-6409.2008.00359.x. ЧВК 2752903. PMID 19789725.
- ^ Мэддисон, Дэвид Р. (11 сентября 2000 г.). «Жесткокрылые. Жук». Веб-проект "Древо жизни". tolweb.org. Получено 18 марта, 2011.
- ^ Нихуис, Оливер; Хартиг, Геррит; Грат, Соня; и другие. (2012). «Геномные и морфологические данные сходятся, чтобы разгадать загадку Strepsiptera». Текущая биология. 22 (14): 1309–1313. Дои:10.1016 / j.cub.2012.05.018. PMID 22704986.
- ^ а б c McKenna, Duane D .; Wild, Александр Л .; и другие. (2015). «Древо жизни жуков показывает, что жесткокрылые пережили массовое вымирание в конце перми, чтобы разнообразить свой вид во время земной революции мелового периода». Систематическая энтомология. 40 (4): 835–880. Дои:10.1111 / syen.12132.
- ^ а б Хант, Тоби; и другие. (2007). «Всеобъемлющая филогения жуков раскрывает эволюционное происхождение сверхизлучения». Наука. 318 (5858): 1913–19116. Bibcode:2007Научный ... 318.1913H. Дои:10.1126 / science.1146954. PMID 18096805. S2CID 19392955.
- ^ «Введение в идентификацию жуков (жесткокрылых)» (PDF). Университет Флориды. Получено 15 марта, 2017.
- ^ Lau T.F.S .; Мейер-Рохов В.Б. (2006). "Половой диморфизм сложного глаза Рагофтальм охбай (Coleoptera; Rhagophthalmidae): I. Морфология и ультраструктура ». Журнал энтомологии Азиатско-Тихоокеанского региона. 9: 19–30. Дои:10.1016 / S1226-8615 (08) 60271-X.
- ^ а б Hangay, G .; Зборовский, П. (2010). Путеводитель по жукам Австралии. CSIRO. п.10.
- ^ Gokan N .; Мейер-Рохов В.Б. (2000). «Морфологические сравнения сложных глаз у Scarabaeoidea (Coleoptera), связанные с максимумами дневной активности жуков и филогенетическими положениями». Журнал сельскохозяйственных наук (Tokyo Nogyo Daigaku). 45 (1): 15–61.
- ^ а б Бениш, Кристоф (2007). «3. Усики, морфология жука». Kerbtier.de (Фауна жуков Германии). Получено 14 марта, 2017.
- ^ Zurek, D.B .; Гилберт, К. (2014). «Статические антенны действуют как локомоторные направляющие, которые компенсируют размытость изображения при дневном зорком хищнике». Труды Королевского общества B: биологические науки. 281 (1779): 20133072. Дои:10.1098 / rspb.2013.3072. ЧВК 3924084. PMID 24500171.
- ^ Берков, Эми; Родригес, Нельсон; Сентено, Педро (2007). «Конвергентная эволюция усиков жука-церамбицида, Onychocerus albitarsis, и укус скорпиона ". Naturwissenschaften. 95 (3): 257–61. Bibcode:2008NW ..... 95..257B. Дои:10.1007 / s00114-007-0316-1. PMID 18004534. S2CID 30226487.
- ^ «Типы антенн». Сиднейский университет. Получено 26 января, 2017.
- ^ Arnett, Jr., R.H .; Томас, М. С. (2001). Американские жуки, том I: Archostemata, Myxophaga, Adephaga, Polyphaga: Staphyliniformia. CRC Press. С. 3–7. ISBN 978-1-4822-7432-5.CS1 maint: несколько имен: список авторов (связь)
- ^ «Анатомия: голова, грудная клетка, брюшко и гениталии (насекомые)». what-when-how.com. Архивировано из оригинал 22 июня 2018 г.. Получено 26 января, 2017.
- ^ Грин, Кристина Карлссон; Ковалев Александр; Свенссон, Эрик I .; Горб, Станислав Н. (2013). «Самец сжимающей способности, женский полиморфизм и сексуальный конфликт: мелкомасштабная морфология надкрылий как сексуально антагонистическая адаптация у самок ныряющих жуков». Журнал интерфейса Королевского общества. 10 (86): 20130409. Дои:10.1098 / rsif.2013.0409. ЧВК 3730688. PMID 23825114.
- ^ Burkness, S .; Хан, Дж. (2007). «Блохи в приусадебных домах». Университет Миннесоты (Расширение). Получено 26 января, 2017.
- ^ а б Карпентер, Джордж Герберт (1899). Насекомые, их строение и жизнь.
- ^ Филипс, Крис; Фред, Элизабет; Кухар, Том, Кожекрылые (Солдатские) Жуки (PDF), заархивировано из оригинал (PDF) 30 ноября 2016 г., получено 14 марта, 2017
- ^ Дональд У. Холл; Марк А. Бранхам (2016). "Calopteron Dispans (Newman) (Insecta: Coleoptera: Lycidae)". Университет Флориды. Получено 15 марта, 2017.
- ^ Lawrence, J. F .; Hastings, A.M .; Dallwitz, M. J .; Пейн, Т. А .; Цурчер, Э. Дж. (2005). "Elateriformia (Coleoptera): описания, иллюстрации, идентификация и поиск информации для семейств и подсемейств ». Получено 26 января, 2017.
- ^ Beenen, R; Винкельман, Дж. К. (2001). «Антекенинген над Chrysomelidae в Нидерландах 5». Entomologische Berichten (на голландском). 61: 63–67.
- ^ Арнетт, Росс Х., младший; Томас, Майкл С. (2000). Американские жуки, том I: Archostemata, Myxophaga, Adephaga, Polyphaga: Staphyliniformia. CRC Press. п. 8. ISBN 978-1-4822-7432-5.
- ^ Сеймур, Роджер С .; Мэтьюз, Филип Г. Д. (2012). «Физические жабры ныряющих насекомых и пауков: теория и эксперимент». Журнал экспериментальной биологии. 216 (2): 164–70. Дои:10.1242 / jeb.070276. PMID 23255190.
- ^ Шмидт-Нильсен, Кнут (15 января 1997 г.). «Дыхание насекомых». Физиология животных: адаптация и окружающая среда (5-е изд.). Издательство Кембриджского университета. п. 55. ISBN 978-0-521-57098-5.
- ^ Миллер, Т.А. (1985). «Глава 8: Строение и физиология кровеносной системы». In Kerkut, G.A .; Гилберт, Л. И. (ред.). Комплексная физиология, биохимия и фармакология насекомых. Том 3: Покровы, дыхание и кровообращение. Pergamom Press. С. 289–355. ISBN 978-0-08-030804-3.
- ^ Скобл, М. Дж. (1992). Чешуекрылые: форма, функции и разнообразие. Издательство Оксфордского университета. ISBN 978-1-4020-6242-1.
- ^ Озбек, Х .; Салокия, Д. (1998). «Вклад в изучение фауны Meloidae (Coleoptera) Турции вместе с новым рекордом». Турецкий зоологический журнал. 22: 23–40.
- ^ Медведев, Л. Н .; Павлов, С. И. (1988). «Брачное поведение Chrysomelidae (Coleoptera)». Энтомологический обзор. 67: 100–108.
- ^ Кроусон, Р. А. (2013). Биология жесткокрылых. Эльзевир Наука. С. 358–370. ISBN 978-1-4832-1760-4.
- ^ Beck, S.D .; Бхарадвадж, Р. К. (1972). «Обратное развитие и клеточное старение у насекомого». Наука. 178 (4066): 1210–1211. Bibcode:1972Научный ... 178.1210Б. Дои:10.1126 / science.178.4066.1210. PMID 4637808. S2CID 34101370.
- ^ "Определение слова" Scarabaeiform "'". Общество энтомологов-любителей. Получено 27 января, 2017.
- ^ Кринский, В. Л. (2009). «8 Жуков (Жесткокрылые) ". В Mullen, G.R .; Durden, L.A. (ред.). Медицинская и ветеринарная энтомология (2-е изд.). Эльзевир. С. 101–115. ISBN 978-0-12-372500-4.
- ^ «Гиперметаморфоз полосатого пузырчатого жука - Epicauta vittata". BugGuide. 2007 г.. Получено 27 января, 2017.
- ^ Цзэн, Юн (1995). «Самый длинный жизненный цикл». Университет Флориды. Получено 17 марта, 2017.
- ^ Джеффрис, Дэниел Л .; Чепмен, Джейсон; Рой, Хелен Э.; Хамфрис, Стюарт; Харрингтон, Ричард; Браун, Питер М. Дж .; Хэндли, Лори-Дж. Лоусон (2013). "Характеристики и движущие силы полета божьих коровок на большой высоте: выводы с помощью вертикального энтомологического радара". PLOS ONE. 8 (2): e82278. Bibcode:2013PLoSO ... 882278J. Дои:10.1371 / journal.pone.0082278. ЧВК 3867359. PMID 24367512.
- ^ Капинера, Джон Л. (2008). Энциклопедия энтомологии (2-е изд.). Springer Science & Business Media. стр.3218–3219. ISBN 978-1-4020-6242-1.
- ^ Arnett R.H., мл .; Томас, М. С. (2001). «Haliplidae». Американские жуки, Том 1. CRC Press. С. 138–143. ISBN 978-0-8493-1925-9.
- ^ «Сосна горная - Жук любви». Парки Канады. Получено 13 марта, 2011.
- ^ а б Мейер-Рохов, В. (1971). «Наблюдения за стридулирующими австралийскими жуками (Hydrophilidae, Cerambycidae, Passalidae, Dynastinae) с использованием сканирующей электронной микроскопии и электрофизиологических методов». Forma et Functio. 4: 326–339.
- ^ Брандмайр П. 1992. Краткий обзор досоциальной эволюции жесткокрылых. Ethol Ecol Evol. 4: 7–16.
- ^ Вятт, Т. Д. и Фостер, В. А. (1989). «Родительская забота о подсоциальном литоральном жуке, Бледиус спектабилис, в отношении паразитизма ихневмонидной осой, Barycnemis blediator". Поведение. 110 (1–4): 76–92. Дои:10.1163 / 156853989X00394. JSTOR 4534785.
- ^ Милн, Лорус Дж .; Милн, Марджери Дж. (1944). «Заметки о поведении погребальных жуков (Nicrophorus spp.)». Журнал Нью-Йоркского энтомологического общества. 52 (4): 311–327. JSTOR 25005075.
- ^ Хански, Илкка; Ив, Камбефор (1991). Экология навозных жуков. Издательство Принстонского университета. С. 626–672. ISBN 978-0-691-08739-9.
- ^ "Жук | Животные и растения зоопарка Сан-Диего". animals.sandiegozoo.org. Получено 18 июля, 2017.
- ^ Chaboo, C.S .; Frieiro-Costa, F.A .; Gómez-Zurita, J .; Вестердайн, Р. (2014). «Субсоциальность листоедов (Coleoptera: Chrysomelidae: Cassidinae, Chrysomelinae)». Журнал естественной истории. 48: 1–44. Дои:10.1080/00222933.2014.909060. S2CID 84683405.
- ^ Чабу, CS (2002). «Первый отчет о неполовозрелых особях, гениталиях и материнском уходе у Eugenysa columbiana (Boheman) (Coleoptera: Chrysomelidae: Cassidinae: Eugenysini)». Бюллетень колеоптерологов. 56: 50–67. Дои:10.1649 / 0010-065x (2002) 056 [0050: froiga] 2.0.co; 2.
- ^ Виндзор, DM (1987). «Естественная история подсоциального жука-черепахи, Acromis sparsa Boheman (Chrysomelidae, Cassidinae) в Панаме». Психея J Энтомол. 94 (1–2): 127–150. Дои:10.1155/1987/19861.
- ^ Рид, САМ; Битсон, М; Hasenpusch, J (2009). «Морфология и биология Pterodunga mirabile Daccordi, необычной субсоциальной хризомелины (Coleoptera: Chrysomelidae)». Дж. Нат Хист. 43 (7–8): 373–398. Дои:10.1080/00222930802586016. S2CID 84744056.
- ^ Виндзор DM, Choe JC. 1994. Истоки родительской заботы у жуков-хризомелид. В: Jolivet PH, Cox ML, Petitipierre E, редакторы. Новые аспекты биологии Chrysomelidae. Серия Entomologica 50. Dordrecht: Kluwer Academic Publishers; п. 111–117.
- ^ Crespi, B.J .; Янега, Д. (1995). «Определение эусоциальности». Поведенческая экология. 6 (1): 109–115. Дои:10.1093 / beheco / 6.1.109.
- ^ а б Кент, Д. С. и Симпсон, Дж. А. (1992). "Эусоциальность жука Austroplatypus incompertus (Coleoptera: Curculionidae) ". Naturwissenschaften. 79 (2): 86–87. Bibcode:1992NW ..... 79 ... 86K. Дои:10.1007 / BF01131810. S2CID 35534268.
- ^ а б «Наука: австралийский жук, который ведет себя как пчела». Новый ученый. 9 мая 1992 г.. Получено 31 октября, 2010.
- ^ Шустер, Джек С .; Шустер, Лаура Б. (1985). «Социальное поведение жуков пассалид (Coleoptera: Passalidae): совместный уход за выводком». Флоридский энтомолог. 68 (2): 266–272. Дои:10.2307/3494359. JSTOR 3494359.
- ^ Вальдес, Хосе В. (2020). «Членистоногие как хищники позвоночных: обзор глобальных моделей». Глобальная экология и биогеография. н / д (н / д). Дои:10.1111 / geb.13157. ISSN 1466-8238.
- ^ Лобанов, А.Л. (2002). "кормление". Биология и экология жуков. Жуки (Coleoptera) и колеоптеролог. Получено 13 марта, 2011.
- ^ Маккой, Питер (2016). Радикальная микология: трактат о наблюдении и работе с грибами. Chthaeus Press. п. 187. ISBN 9780986399602.
- ^ Ислам, М .; Hossain, A .; Мостафа, М. Г .; Хоссейн, М. М. (2016). «Важные с судебной точки зрения насекомые, связанные с разложением мышей падалью в Бангладеш». Журнал биологических наук Джахангирнагарского университета. 5 (1): 11–20. Дои:10.3329 / jujbs.v5i1.29739.
- ^ Гребенников, Василий В .; Лешен, Ричард А. Б. (2010). «Наружные полости экзоскелета у Coleoptera и их возможные микангиальные функции». Энтомологическая наука. 13 (1): 81–98. Дои:10.1111 / j.1479-8298.2009.00351.x.
- ^ а б c Эванс и Беллами (2000), стр. 27–28
- ^ Котт, Х. (1940). Адаптивная окраска животных. Метуэн. п.414.
- ^ а б c Эванс и Беллами (2000), п. 126
- ^ а б Эванс и Беллами (2000)
- ^ МакХью (2009), п. 199
- ^ Пек, Стюарт Б. (2006). «Распространение и биология эктопаразитического бобрового жука. Platypsyllus castoris Рицема в Северной Америке (Coleoptera: Leiodidae: Platypsyllinae) ». Insecta Mundi. 20 (1–2): 85–94.
- ^ Нойман, П., Эльзен, П. Дж. (2004). «Биология мелкого ульевика (Aethina tumida, Coleoptera: Nitidulidae): пробелы в наших знаниях об инвазивных видах ». Apidologie. 35 (3): 229–247. Дои:10.1051 / apido: 2004010.
- ^ Мейер, Джон Р. (8 марта 2005 г.). «Жесткокрылые». Университет штата Северная Каролина. Архивировано из оригинал 24 мая 2000 г.. Получено 13 марта, 2011.
- ^ Вебер, Дональд С .; Саска, Павел; Чабу, Кэролайн С. (2008). «Жуки-жуки (Coleoptera: Carabidae) как паразитоиды». Энциклопедия энтомологии. Springer Нидерланды. С. 719–721. Дои:10.1007/978-1-4020-6359-6_492. ISBN 978-1-4020-6242-1.
- ^ Gullan, P.J .; Крэнстон, П. С. (2014). Насекомые: очерк энтомологии (5-е изд.). Wiley, John & Sons. п. 314. ISBN 978-1-4443-3036-6.
- ^ Джонс, Г. Д. и Джонс, С. Д. (2001). «Использование пыльцы и ее значение для энтомологии». Неотропическая энтомология. 30 (3): 314–349. Дои:10.1590 / S1519-566X2001000300001.
- ^ Ollerton, J .; Johnson, S.D .; Кранмер, Л. и Келли, С. (2003). «Экология опыления скопления луговых асклепиад в Южной Африке». Анналы ботаники. 92 (6): 807–834. Дои:10.1093 / aob / mcg206. ЧВК 4243623. PMID 14612378.
- ^ а б Malloch, D .; Блэквелл, М. (1993). «Дисперсионная биология офиостоматоидных грибов». В Вингфилде, М. Дж .; К. А. Зайферт; Дж. Ф. Уэббер (ред.). Ceratocystis и Ophiostoma: систематика, экология и патология. Святой Павел: APS. С. 195–206. ISBN 978-0-89054-156-2.
- ^ Скотт, Дж. Дж .; О, Д. С .; Yuceer, M. C .; Klepzig, K. D .; Clardy, J .; Карри, К. Р. (2008). «Бактериальная защита жуков-грибов мутуализма». Наука. 322 (5898): 63. Bibcode:2008Научный ... 322 ... 63S. Дои:10.1126 / наука.1160423. ЧВК 2761720. PMID 18832638.
- ^ Франк-Гроссманн, Х. (1967). «Эктосимбиоз у насекомых, населяющих древесину». В М. Генри (ред.). Симбиоз. 2. Нью-Йорк: Академическая пресса. С. 141–205.
- ^ Ходек, Иво (2012). «Обзорная статья: Диапауза взрослых особей жесткокрылых». Психея: журнал энтомологии. 2012: 1–10. Дои:10.1155/2012/249081.
- ^ Кауфманн, Т. (1971). «Зимовка арктического жука, Pterostichus brevicornis, на Аляске ». Журнал Канзасского энтомологического общества. 44 (1): 81–92.
- ^ Gullan, P.J .; Крэнстон, П. С. (1994). Насекомые: очерк энтомологии. Чепмен и Холл. С. 103–104. ISBN 978-0-412-49360-7.
- ^ Ломбадеро, Мария Дж .; Эйрес, Мэтью П .; Эйрес, Брюс Д .; Рив, Джон Д. (2000). «Холодоустойчивость четырех видов короеда (Coleoptera: Scolytidae) в Северной Америке» (PDF). Экология окружающей среды. 29 (3): 421–432.
- ^ Сформо, Т .; Уолтерс, К .; Jeannet, K .; Wowk, B .; Fahy, G.M .; Barnes, B.M .; Думан, Дж. Г. (2010). «Глубокое переохлаждение, остекловывание и ограниченная выживаемость до −100 ° C у аляскинского жука. Cucujus clavipes puniceus (Coleoptera: Cucujidae) личинки ". Журнал экспериментальной биологии. 213 (3): 502–509. Дои:10.1242 / jeb.035758. PMID 20086136.
- ^ Брукс, Кристофер (26 марта 2013 г.). «Жизнь экстремофилов: выживание во враждебных средах обитания». BBC Nature. Получено 16 марта, 2017.
- ^ Graham, L.A; Liou, Y.C .; Уокер, В. К .; Дэвис, П. Л. (август 1997 г.). «Гиперактивный протеин-антифриз от жуков». Природа. 388 (6644): 727–728. Bibcode:1997Натура.388..727Г. Дои:10.1038/41908. PMID 9285581. S2CID 205029622.
Желтый мучной червь, Тенебрио Молитор, содержит семейство небольших белков термического гистерезиса, богатых Cys и Thr, которые понижают точку замерзания гемолимфы ниже точки плавления на целых 5,58 ° C (ΔT = тепловой гистерезис). Экспрессию белков теплового гистерезиса оценивали на протяжении всего развития и после воздействия измененных условий окружающей среды.
- ^ Уолтерс, К. Р. младший; Серианни, А. С .; Сформо, Т .; Barnes, B.M .; Думан, Дж. Г. (2009). «Небелковый антифриз ксиломаннана, вызывающий термический гистерезис, у морозостойкого аляскинского жука Upis ceramboides». PNAS. 106 (48): 20210–20215. Bibcode:2009PNAS..10620210W. Дои:10.1073 / pnas.0909872106. ЧВК 2787118. PMID 19934038.
- ^ Уолтерс младший, К. Р .; Pan, Q .; Серианни, А. С .; Думан, Дж. Г. (2009). «Биосинтез криопротекторов и избирательное накопление треитола у морозостойкого аляскинского жука, Upis ceramboides". Журнал биологической химии. 284 (25): 16822–16831. Дои:10.1074 / jbc.M109.013870. ЧВК 2719318. PMID 19403530.CS1 maint: несколько имен: список авторов (связь)
- ^ Эдни, Э. Б. (1971). «Температура тела жуков-тенебрионид в пустыне Намиб на юге Африки» (PDF). Журнал экспериментальной биологии. 55: 253–272.
- ^ Knisley, C.B .; Schultz, T. D .; Хазевинкель, Т. Х. (1990). «Сезонная активность и терморегуляторное поведение Cicindela patruela (Coleoptera: Cicindelidae) ". Анналы Энтомологического общества Америки. 83 (5): 911–915. Дои:10.1093 / aesa / 83.5.911.
- ^ Паркер, Эндрю Р .; Лоуренс, Крис Р. (1 ноября 2001 г.). «Захват воды пустынным жуком». Природа. 414 (6859): 33–34. Bibcode:2001Натура.414 ... 33П. Дои:10.1038/35102108. PMID 11689930. S2CID 34785113.
- ^ Хобак, У. Вятт; Стэнли, Дэвид В .; Хигли, Леон G .; Барнхарт, М. Кристофер (1998). «Выживание личинок тигровых жуков в условиях погружения и аноксии. Cicindela togata". Американский натуралист из Мидленда. 140 (1): 27–33. Дои:10.1674 / 0003-0031 (1998) 140 [0027: SOIAAB] 2.0.CO; 2.
- ^ Конради-Ларсен, Эльза-Маргрет; Сёмме, Лауриц (1973). «Анаэробиоз у зимующих жуков. Pelophila borealis". Природа. 245 (5425): 388–390. Bibcode:1973Натура.245..388С. Дои:10.1038 / 245388a0. S2CID 4288059.
- ^ Эллисон, Р. (17 марта 2015 г.). «На половине территории Великобритании начинается миграция пыльцевых жуков». Получено 16 марта, 2017.
- ^ Саутвуд, Т. Р. Э. (1962). «Миграция наземных членистоногих относительно среды обитания». Биологические обзоры. 37 (2): 171–211. Дои:10.1111 / j.1469-185X.1962.tb01609.x.
- ^ Дингл, Х. (2014). Миграция: биология жизни в движении. Издательство Оксфордского университета.
- ^ а б Заблудов, Марк (2008). Жуки. Малайзия: Мишель Бизон. С. 14–17. ISBN 978-0-7614-2532-8.
- ^ Cambefort, Ив (2011). «Жуки как религиозные символы». Insects.org. Архивировано из оригинал 18 октября 2016 г.. Получено 10 февраля, 2017.
- ^ Доллингер, Андре (январь 2002 г.). «Древнеегипетский бестиарий: Насекомые». Архивировано из оригинал 1 апреля 2015 г.. Получено 19 июля, 2011.
- ^ Моралес-Корреа, Бен (2006). «Египетские символы». Все-о-Египте. Архивировано из оригинал 20 августа 2011 г.. Получено 19 июля, 2011.
- ^ Уссишкин, Давид (2004). Новые археологические раскопки в Лахисе (1973–1994). Тель-Авив: Институт археологии Тель-авивский университет. Архивировано из оригинал 3 июля 2018 г.. Получено 28 декабря, 2015.
- ^ "Исида и Осирис", Мораль, в томе V Классическая библиотека Леба, 1936. «Раса жуков не имеет самок, но все самцы выбрасывают свою сперму в круглую гранулу материала, которую они сворачивают, толкая ее с противоположной стороны, так же, как кажется, что солнце поворачивает небеса в противоположном направлении. по своему собственному курсу, который идет с запада на восток.
- ^ Бец, Х., изд. (1992). Греческий магический папирус в переводе (включая демотические заклинания) (2-е изд.). Издательство Чикагского университета. С. IV.52–85, VII.520, XII.101, XIII.1065–1070, XXXVI.170.
- ^ Плиний Старший. "Естественная история Книга 11 ". Perseus.tufts.edu. Получено 25 января, 2017.
- ^ Плиний Старший. "Естественная история Книга 11, глава 34 ". Perseus.tufts.edu. Получено 25 января, 2017.
- ^ а б Бивис, И. К. (1988). Насекомые и другие беспозвоночные в классической древности. Эксетерский университет. С. 153–154.
- ^ Антонин Либералис. Метаморфозы. Пер. Селория, Ф. 1992. Метаморфозы Антонина Либералиса: перевод с комментарием. Лондон и Нью-Йорк, Рутледж.
- ^ Sprecher-Uebersax, E. (2008). «Жук-олень Lucanus Cervus (Coleoptera, Lucanidae) в искусстве и мифологии». Revue d'Ecologie. 63: 145–151.
- ^ а б Государственный университет Миссисипи. «История долгоносика в США». Экономические последствия коробочного долгоносика. Архивировано из оригинал 12 мая 2008 г.
- ^ Allen, E.A .; Хамбл, Л. М. (2002). «Интродукция неместных видов: угроза лесам и лесной экономике Канады». Канадский журнал патологии растений. 24 (2): 103–110. Дои:10.1080/07060660309506983. S2CID 85073955.
- ^ Уэббер, Джоан Ф. (2000). "Поведение насекомых-переносчиков и эволюция болезни голландского вяза". Вязы. п. 47. Дои:10.1007/978-1-4615-4507-1_3. ISBN 978-1-4613-7032-1.
- ^ Алехин, А .; Baker, M .; Mota-Sanchez, D .; Dively, G .; Графиус, Э. (2008). «Устойчивость колорадского жука к инсектицидам». Американский журнал исследований картофеля. 85 (6): 395–413. Дои:10.1007 / s12230-008-9052-0. S2CID 41206911.
- ^ Локвуд, Джеффри А. (21 октября 2007 г.). «Баг-бомба: почему наш следующий теракт может произойти на шести ногах». Бостон Глобус. Получено 13 февраля, 2017.
- ^ Хизер, Нил У .; Холлман, Гай Дж. (2008). Борьба с вредителями и фитосанитарные торговые барьеры. КАБИ. стр.17–18. Дои:10.1079/9781845933432.0000. ISBN 978-1-84593-343-2.
- ^ Адкок, Эдвард (2005). «Вредители - Жук-дозор». Сохранение и коллективный уход. Оксфордский университет. Архивировано из оригинал 10 июля 2011 г.. Получено 17 июля, 2011.
- ^ Такано, Сюн-Ичиро; Такасу, Кейджи; Фусими, Цутому; Ichiki, Ryoko T .; Накамура, Сатоши (2014). «Пригодность четырех видов пальм для развития инвазивных вредителей. Бронтиспа longissima (Coleoptera: Chrysomelidae) в поле ". Энтомологическая наука. 17 (2): 265–268. Дои:10.1111 / ens.12048.
- ^ "Горный сосновый жук в Британской Колумбии". Природные ресурсы Канады. 19 августа 2008 г. Архивировано с оригинал 19 апреля 2010 г.. Получено 24 июня, 2010.
- ^ «Насекомые: Coleoptera: Coccinellidae». Институт продовольственных и сельскохозяйственных услуг, Университет Флориды. 2014. Получено 12 февраля, 2017.
- ^ "'В Великобритании обнаружена смертельная божья коровка ». Новости BBC. 5 октября 2004 г.. Получено 17 июня, 2010.
- ^ Кромп, Б. (1999). «Жужелицы в устойчивом сельском хозяйстве: обзор эффективности борьбы с вредителями, аспектов выращивания и улучшения». Сельское хозяйство, экосистемы и окружающая среда. 74 (1–3): 187–228. Дои:10.1016 / S0167-8809 (99) 00037-7.
- ^ «Жуки играют важную роль в борьбе с сорняками». Rothamsted Research. Получено 14 марта, 2017.
- ^ «Zygogramma bicolorata (мексиканский жук)». Компендиум инвазивных видов. 2012 г.. Получено 12 февраля, 2017.
- ^ Ковалев, О. В .; Резник, С.Ю .; Черкашин, В. Н. (1983). «Особенности методов использования Zygogramma Chevr. (Coleoptera, Chrysomelidae) в биологической борьбе с амброзией (Ambrosia artemisiifolia L., A. psilostachya D.C.)». Энтомологическое обозрение (на русском). 62: 402–408.
- ^ а б Борнемесса, Джордж (1970). "Исследования насекомых по борьбе с навозными мухами с помощью навозных жуков", Онтофаг Газелла F. (Coleoptera: Scarabaeinae) ". Австралийский журнал энтомологии. 9: 31–41. Дои:10.1111 / j.1440-6055.1970.tb00767.x.
- ^ Браун, Жаклин; Scholtz, Clarke H .; Жано, Жан-Луи; Греллье, Серафина и Подвоевски, Паскаль (2010). «Навозные жуки (Coleoptera: Scarabaeidae) могут улучшить гидрологические свойства почвы». Прикладная экология почвы. 46 (1): 9–16. Дои:10.1016 / j.apsoil.2010.05.010. HDL:2263/14419.
- ^ Лоузи, Джон Э .; Воан, Мейс (2006). «Экономическая ценность экологических услуг, предоставляемых насекомыми» (PDF). Бионаука. 56 (4): 311–323. Дои:10.1641 / 0006-3568 (2006) 56 [311: TEVOES] 2.0.CO; 2. Архивировано из оригинал (PDF) 19 июня 2018 г.. Получено 17 июля, 2011.
- ^ Томберлин, Джеффри К .; Сэнфорд, Мишель Р. (2012). «Судебная энтомология и животный мир». У Хаффмана, Джейн Э .; Уоллес, Джон Р. (ред.). Криминалистика дикой природы: методы и приложения. Развитие судебной медицины. 6 (2-е изд.). Джон Уайли и сыновья. С. 81–107. ISBN 978-1-119-95429-3.
- ^ Жуки-дерместиды работают над черепом самки черного медведя. 2014. Получено 26 января, 2017.
- ^ Фернандес-Ялво, Иоланда; Монфорт, Мария Долорес Марин (2008). «Экспериментальная тафономия в музеях: протоколы подготовки скелетов и ископаемых позвоночных животных под растровой электронной микроскопией». Geobios. 41: 157–181. Дои:10.1016 / j.geobios.2006.06.006.
- ^ Рамос-Элордуй, Джульета; Мензель, Питер (1998). Жуткая кухня: путеводитель по съедобным насекомым для гурманов. Внутренние традиции / Медведь и Компания. п. 5. ISBN 978-0-89281-747-4.
- ^ Холланд, Дженнифер С. (14 мая 2013 г.). «ООН призывает к поеданию насекомых; 8 популярных ошибок, которые стоит попробовать». Национальная география. Получено 26 января, 2017.
- ^ Мейер-Рохов, В. Б. (январь 2017 г.). «Лечебные членистоногие и другие, в основном наземные, важные в народной медицине беспозвоночные: сравнительный обзор и обзор». Журнал этнобиологии и этномедицины. 13 (9): 9. Дои:10.1186 / s13002-017-0136-0. ЧВК 5296966. PMID 28173820.
- ^ Пирсон, Дэвид Л .; Кассола, Фабио (1992). «Модели видового богатства тигровых жуков (Coleoptera: Cicindelidae) во всем мире: индикаторный таксон для исследований в области биоразнообразия и сохранения». Биология сохранения. 6 (3): 376–391. Дои:10.1046 / j.1523-1739.1992.06030376.x. JSTOR 2386038.
- ^ МакГеоч, Мелоди А .; Ван Ренсбург, Берндт Дж .; Ботес, Антуанетта (2002). «Проверка и применение биоиндикаторов: тематическое исследование навозных жуков в экосистеме саванны». Журнал прикладной экологии. 39 (4): 661–672. Дои:10.1046 / j.1365-2664.2002.00743.x.
- ^ Лаша, Тибо; Вермелингер, Бит; Госснер, Мартин М .; Бусслер, Хайнц; Исакссон, Гуннар; Мюллер, Йорг (2012). «Сапроксилированные жуки как виды-индикаторы количества валежной древесины и температуры в буковых лесах Европы». Экологические показатели. 23: 323–331. Дои:10.1016 / j.ecolind.2012.04.013.
- ^ Бохач, Ярослав (1999). «Жуки-стафилиниды как биоиндикаторы» (PDF). Сельское хозяйство, экосистемы и окружающая среда. 74 (1–3): 357–372. CiteSeerX 10.1.1.496.4273. Дои:10.1016 / S0167-8809 (99) 00043-2.
- ^ Жизненный цикл самоцветов округлой формы Sternocera spp. วงจร ชีวิต ของ แมลง ทับ กลม ใช้ เวลา นาน ถึง 2 ปี - Сиамский зоопарк насекомых. Malaeng.com (20 октября 2008 г.). Проверено 4 апреля 2013.
- ^ Айви, Майкл А. (2002). «105. Zopheridae». В Росс Х. Арнетт; Майкл Чарльз Томас (ред.). Американские жуки: Polyphaga: Scarabaeoidea через Curculionoidea. Том 2 американских жуков. CRC Press. стр.457–462. ISBN 978-0-8493-0954-0.
- ^ Реннессон, Стефан; Сезар, Николас; Гримо, Эммануэль (2008). "Duels en miniature: la délicate mise en scène des scarabées au nord de la Thaïlande" (PDF). Насекомые (На французском). 3 (151).
- ^ а б Эральдо Медейрос Коста-Нето (2003). «Развлечения с насекомыми: песни и борьба с насекомыми по всему миру. Краткий обзор» (PDF). Etnobiología. 3: 21–29. Архивировано из оригинал (PDF) 13 февраля 2017 г.. Получено 12 февраля, 2017.
- ^ Пембертон, Р. В. (1990). «Корейская игра с водяными жуками». Пан-Тихоокеанский энтомолог. 66 (2): 172–174.
- ^ Bateman, C .; Халкр, Дж. (2016). «Хищные ныряющие жуки как домашние животные и самоочищающийся аквариум» (PDF). Университет Флориды (Продление МФСА). Получено 27 января, 2017.
- ^ Кавахара, А. Ю. (2007). «Тридцатифутовые телескопические сети, видеоигры для сбора насекомых и домашние жуки: энтомология в современной Японии» (PDF). Американский энтомолог. 53 (3): 160–172. Дои:10.1093 / ae / 53.3.160. Архивировано из оригинал (PDF) 12 апреля 2016 г.. Получено 27 января, 2017.
- ^ Мэй, Митчелл (11 июля 1999 г.). «Йена за ошибки». Чикаго Трибьюн. Получено 27 января, 2017.
- ^ Уоттс, Джонатан (11 августа 1999 г.). «Торговец жуками». Хранитель. Получено 27 января, 2017.
- ^ Морс, Дебора Дененхольц; Данахай, Мартин А. (2007). Викторианские сны о животных: изображения животных в викторианской литературе и культуре. Издательство Ashgate. п. 5. ISBN 978-0-7546-5511-4.
Викторианская мания собирать жуков
- ^ Уоллес, Альфред Рассел (1869). Малайский архипелаг: земля орангутанов и райских птиц. Повествование о путешествии с зарисовками человека и природы. (1-е изд.). Макмиллан. стр. vii – xiv.
- ^ Шведская биомиметика: технология платформы μMist (исходный URL = http://www.swedishbiomimetics.com/biomimetics_folder.pdf) (дата архивации = 13 декабря 2013 г.)
- ^ «Намибский пустынный жук вдохновляет самонаполняющуюся бутылку с водой». Новости BBC. 23 ноября 2012 г.
- ^ Певица Эмили (29 января 2009 г.). "Армейский жук с дистанционным управлением". Обзор технологий MIT. Получено 16 марта, 2017.
- ^ Цао, Фэн; Чжан, Чао; Хао Ю Чу; Сато, Хиротака (2016). «Робот-гибрид насекомых и компьютеров с регулируемыми пользователем скоростью, длиной шага и походкой». Журнал интерфейса Королевского общества. 13 (116): 20160060. Дои:10.1098 / rsif.2016.0060. ЧВК 4843679. PMID 27030043.
- ^ Сато, Хиротака; Доан, Тат Тханг Во; Колев, Святослав; Huynh, Ngoc Anh; Чжан, Чао; Massey, Travis L .; Клиф, Джошуа ван; Икеда, Кадзуо; Аббель, Питер (16 марта 2015 г.). «Расшифровка роли управляющей мышцы жесткокрылых с помощью стимуляции свободного полета». Текущая биология. 25 (6): 798–803. Дои:10.1016 / j.cub.2015.01.051. PMID 25784033.
- ^ Во Доан, Тат Тханг; Tan, Melvin Y.W .; Буй, Сюань Хиен; Сато, Хиротака (3 ноября 2017 г.). «Сверхлегкий и живоногий робот». Мягкая робототехника. 5 (1): 17–23. Дои:10.1089 / soro.2017.0038. PMID 29412086.
- ^ Викрам Айер1; Али Наджафи; Йоханнес Джеймс; Сойер Фуллер; Шьямнатх Голлакота (июль 2020 г.). «Беспроводное управляемое видение для живых насекомых и роботов-насекомых». Научная робототехника. 5 (44): eabb0839. Дои:10.1126 / scirobotics.abb0839. S2CID 220688078.
- ^ Роб Пичета (15 июля 2020 г.). «Ученые прикрепили к жуку крошечную камеру, чтобы проверить, насколько маленькими могут быть видеотехнологии». CNN.
- ^ Коце, Д. Йохан; О'Хара, Роберт Б. (2003). «Виды сокращаются - но почему? Объяснение сокращения численности жужелиц (Coleoptera, Carabidae) в Европе». Oecologia. 135 (1): 138–148. Bibcode:2003Oecol.135..138K. Дои:10.1007 / s00442-002-1174-3. PMID 12647113. S2CID 11692514.
- ^ Хански, Илкка; Койвулехто, Елена; Кэмерон, Элисон; Рахагалала, Пьер (2007). «Вырубка лесов и очевидное исчезновение эндемичных лесных жуков на Мадагаскаре». Письма о биологии. 3 (3): 344–347. Дои:10.1098 / рсбл.2007.0043. ЧВК 1995085. PMID 17341451.
- ^ Campanaro, A .; Zapponi, L .; Hardersen, S .; Méndez, M .; Al Fulaij, N .; Audisio, P .; Бардиани, М .; Карпането, Г. М .; Corezzola, S .; Della Rocca, F .; Harvey, D .; Hawes, C .; Kadej, M .; Karg, J .; Каток, М .; Смолис, А .; Sprecher, E .; Thomaes, A .; Toni, I .; Vrezec, A .; Заули, А .; Зилиоли, М .; Киари, С. (2016). «Европейский протокол мониторинга жука-оленя, флагманского сапроксильного вида». Водолазы-консерваторы. 9 (6): 574–584. Дои:10.1111 / icad.12194.
- ^ Новый, Т. Р. (2009). Жуки в сохранении. Джон Вили и сыновья. pp. ix, 1–26, passim. ISBN 978-1-4443-1863-0.
Библиография
- Эванс, Артур V .; Беллами, Чарльз (2000). Чрезмерная любовь к жукам. Калифорнийский университет Press. ISBN 978-0-520-22323-3.
- МакХью, Джозеф V .; Либхерр, Джеймс К. (2009). «Жесткокрылые». В Винсенте Х. Реше; Кольцо Т. Карде (ред.). Энциклопедия насекомых (2-е изд.). Академическая пресса. стр.183–201. ISBN 978-0-12-374144-8.
дальнейшее чтение
- Бекманн, Пол (2001). Живые драгоценности: естественный дизайн жуков. ISBN 978-3-7913-2528-6.
- Cooter, J .; М. В. Л. Барклай, ред. (2006). Справочник колеоптеролога. Любительское энтомологическое общество. ISBN 978-0-900054-70-9.
- Жуки-личинки мира. Энтомологическое общество Америки. ISBN 978-0-643-05506-3.
- Гримальди, Дэвид; Майкл С. Энгель (16 мая 2005 г.). Эволюция насекомых. ISBN 978-0-521-82149-0.
- Харде, К. В. (1984). Полевой справочник по окраске жуков. С. 7–24. ISBN 978-0-7064-1937-5.
- Уайт, Р. Э. (1983). Жуки. Хоутон Миффлин. ISBN 978-0-395-91089-4.
внешняя ссылка
| Викивиды имеет информацию, относящуюся к Жесткокрылые |
| Викибук Дихотомический ключ есть страница по теме: Жесткокрылые |
- Жесткокрылые от Веб-проект "Древо жизни"
- (на немецком) Кефер дер Вельт
- Coleoptera Atlas
- Жуки - Жесткокрылые
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
